
A just-released report on the latest count for the Alaska portion of the Southern Beaufort subpopulation reveals that numbers have been stable since 2010 despite claims the population has continued to decline. However, the study also has a very odd feature: 2012 had the highest population estimate over the decade of 2006-2015 yet also had the lowest survival of all age classes since 2001.

Healthy polar bear male at Kaktovik, Alaska on the Southern Beaufort Sea, September 2019, Ed Boudreau photo, with permission.
However, what is essentially good news about polar bear health and survival in the Southern Beaufort has so far been glossed over by the media because the report prominently includes estimates of polar bear dens on land in areas of potential oil exploration, a highly politicized topic. Accordingly, the Washington Post (picked up by other outlets) focused a statement in the paper that “long-term persistence of polar bears (Ursus maritimus) is threatened by sea-ice loss due to climate change” and on the denning issue rather than the new population count. As far as I am aware, no other population estimate report has included such distracting information.
Recent claim of a polar bear expert [my bold]:
In 2015…the polar bear population in the Beaufort Sea had declined by 40% over the previous decade. “We can only anticipate that those declines have continued.” Steven Amstrup, 29 September 2019.
Area of the study 
Population count
For the last five years, the Southern Beaufort subpopulation has been the jewel in the crown of those who promote the notion that polar bears are threatened with extinction due to man-made global warming. With an estimated decline of 25-50% between 2001 and 2010 (often ‘simplified’ as “40%”, see quote above), it has been the most dramatic example of a population decline blamed on lack of summer sea ice – even though it is known that the decline was actually the result of thick spring ice conditions in the period 2004-2006 and that Inuit in the Canadian portion of the Southern Beaufort disagree with this assessment (Crockford 2019, 2020).
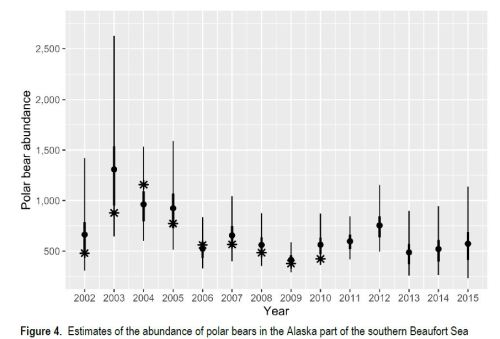
The last population count of polar bears in the Southern Beaufort Sea covered the years 2001-2010 (Bromaghin et al. 2015). In that study, the 2010 estimate for the same area of Alaska as the most recent study was 562 (range, 363-873). For 2015 , the estimate was 573 (range, 232-1,140) – see their Fig. 4 below – and the average estimated abundance for the entire 2006-2015 period was 563 (range, 340-920)(Atwood et al. 2020).
In other words, no change.
However, not mentioned in the paper but obvious in the graph below is that the population count in 2012 was the highest over the 2006-2015 period – an average of about 750 bears.
Survival estimates by age

Oddly, all age classes (including adults) had extremely low apparent survival rates in 2012 compared to all other years since 2001: not even the known ‘poor survival’ years of 2004-2006 shown in the graphs were as bad (see also Bromaghin et al. 2015; Stirling et al. 2008). Even more peculiar is the fact that no suggestions whatsoever are offered by the authors to explain this phenomenon: it is simply dismissed as a fluctuation. Similarly, nothing is said about the surprising fact that the population estimate for that year was the highest over the ten year period of 2006-2015 (see section above).
Was the apparent low survival in 2012 a result of the failure to account for movement of bears? This seems likely, as the authors don’t attempt to blame this on lack of summer sea ice, which would be the predictable response. If there had been a negative impact of much reduced summer sea ice or even thick spring ice on Southern Beaufort bears in 2012, it wasn’t evident by the fall of 2012, when a US Fish and Wildlife Service survey found polar bear numbers higher than they had been in a decade (USFWS 2013:17). Neither were starving bears or reduced numbers of cubs evident by 2013, when researchers were doing field research in the area (Rode et al 2013, 2014, 2018).
Movement of bears
Perhaps critically, this study model was not able to adequately account for the fact that Alaskan polar bears move around: they are known to move to and from nearshore and offshore locations (see map of study area above), as well as to and from neighbouring subpopulations (see also next section), which may have affected the accuracy of the population and survival estimates. For example:
“Analyses of relocations of polar bears carrying satellite radio collars suggested that at Utqiağvik (formerly Barrow), Alaska, in the west, 50% of polar bears were from the SB subpopulation and 50% were from the Chukchi Sea (CS) subpopulation.” PBSG.
This shortcoming was also present in the model used to generate the last population estimate but the potential effect was dismissed as “likely to be small” (Bromaghin et al. 2015:646). However, rather than do the work necessary to resolve this potentially critical problem, USGS authors went ahead and generated another population estimate and more survival estimates using a model with a known flaw. We know this because the authors (Atwood et al. 2020:12) state: “Additional work to investigate the influence of bear movement on abundance estimates is underway.”
Beaufort Sea thick spring ice and movement of polar bears: a reminder
Ian Stirling (2002:68) had this to say about the reduced survival of polar bears due thick spring ice in the Beaufort Sea, that could also be said to apply to the similar events that took place in 2004-2006:
“In the eastern Beaufort Sea, in years during and following heavy ice conditions in spring, we found a marked reduction in production of ringed seal pups and consequently in the natality of polar bears (Stirling and Lunn, 1997). The effect appeared to last for about three years, after which productivity of both seals and bears increased again. These clear and major reductions in productivity of ringed seals in relation to ice conditions occurred at decadal scale intervals in the mid-1970s and 1980s (Fig. 5) and, on the basis of less complete data, probably in the mid-1960s as well (Stirling et al., 1977b; Stirling and Lunn, 1997).” [my bold]
Below is an excerpt from a previous post on an incident of cannibalism that occurred in 1976 that’s pertinent because of the descriptions of the spring ice conditions for 1975 and 1976, and the effects these had on both ringed seals and polar bears. A report of an incident of cannibalism in 1976 was made by biologist Jack Lentfer in his summary of polar bear research in Alaska for the IUCN Polar Bear Specialist Group’s 6th meeting (1976, pg. 187 and 192, “Polar bear management and research in Alaska 1974-76”). See the map below for locations of communities mentioned:
“Both 1975 and 1976 were “heavy” ice years. Bears traveled south to the southern Chukchi Sea and northern Bering Sea with a movement of heavy ice south early in the winter of these years. Unusually high kills [of polar bears] occurred on St. Lawrence Island and near the villages of Wales and Shishmaref in areas where bears are seldom encountered.
…
Sea ice was heavier than usual and heavy ice extended further south than usual in 1975 and 1976. This may have caused a movement of ringed seals, the principal food of bears, to the west and south (Burns et al. 1976) and a similar movement of some bears. In both years significantly fewer seals were killed by bears in the Barrow, Alaska area than in years when ice had not been so heavy. Also, perhaps related to reduced availability of seals, the first instance of predation by an adult bear, a male, on cubs was noted.
Both cubs in a litter were nearly completely consumed on 13 April 1976.” [my bold]
Seal biologists John Burns and colleagues described the unusual nature of the movements of seals and bears in their quarterly report on research they were doing on the distribution and abundance of seals in 1974 and 1975 (Burns et al. 1975). They say (in part):
“Survey results substantiated that a major, short-term shift in density had indeed occurred. Density of seals in the Beaufort Sea was down 10 fold with a corresponding increase in the Chukchi Sea of between 10 and 15 fold.
…In my opinion, the overriding factor affecting ringed seal distribution is the distribution of favorable sea ice conditions. From past experience it was obvious the prevailing sea ice conditions in the Beaufort Sea were, by and large, unfavorable for breeding ringed seals whereas they were excellent in the Chukchi Sea.
…The distribution of ringed seals directly affected the distribution of their most significant predator, the polar bear.” [my bold]
 As Burns and colleagues pointed out, the ice conditions of 1974-1976 in Alaska and western Canada were unusual. In their 2008 paper on Eastern Beaufort polar bears, Ian Stirling and colleagues said that prior to 2004-2006 (when there was heavy spring ice and some very thin bears), they had not seen similar development of shorefast pressure ridges since 1974 — even though they had worked there from 1971 to 1979 and from 1985 to 1987. This suggests that conditions in the eastern Beaufort were worse in the spring of 1974 (with almost as bad conditions the following two years, see previous post here), but that in the Chukchi Sea and western Beaufort, the worst effects were evident in 1975 and 1976.
As Burns and colleagues pointed out, the ice conditions of 1974-1976 in Alaska and western Canada were unusual. In their 2008 paper on Eastern Beaufort polar bears, Ian Stirling and colleagues said that prior to 2004-2006 (when there was heavy spring ice and some very thin bears), they had not seen similar development of shorefast pressure ridges since 1974 — even though they had worked there from 1971 to 1979 and from 1985 to 1987. This suggests that conditions in the eastern Beaufort were worse in the spring of 1974 (with almost as bad conditions the following two years, see previous post here), but that in the Chukchi Sea and western Beaufort, the worst effects were evident in 1975 and 1976.
References
Atwood, T.C., Bromaghin, J.F., Patil, V.P., Durner, G.M., Douglas, D.C., and Simac, K.S., 2020. Analyses on subpopulation abundance and annual number of maternal dens for the U.S. Fish and Wildlife Service on polar bears (Ursus maritimus) in the southern Beaufort Sea, Alaska: U.S. Geological Survey Open-File Report 2020-1087. https://doi.org/10.3133/ofr20201087. pdf here.Abstract The long-term persistence of polar bears (Ursus maritimus) is threatened by sea-ice loss due to climate change, which is concurrently providing an opportunity in the Arctic for increased anthropogenic activities including natural resource extraction. Mitigating the risk of those activities, which can adversely affect the population dynamics of the southern Beaufort Sea (SBS) subpopulation, is an emerging challenge as polar bears become more reliant on land and come into more frequent contact with humans. The Marine Mammal Protection Act and Endangered Species Act require the U.S. Fish and Wildlife Service to determine whether industrial activities will have a negligible impact on the SBS subpopulation. Information important to making that determination includes estimates of subpopulation abundance and the number of maternal dens likely to be present in areas where industrial activities occur. We analyzed mark-recapture data collected from SBS polar bears sampled in Alaska during 2001–16 using multistate Cormack-Jolly-Seber models. Estimated survival rates were relatively high during 2001–03, lower during 2004–08, then higher during 2009–15 except for 2012. Estimated abundance in the Alaska part of the SBS was consistent with the estimated survival rates, declining from about 1,300 bears in 2003 to 525 bears in 2006 and then remaining generally stable during 2006–15. The point estimate for the Alaska part of the SBS in 2015, the last year in which abundance could be estimated, was 573 bears (95-percent credible interval = 232, 1,140 bears). To estimate the expected number of terrestrial dens likely to be present in a given region in a given year, we used a Bayesian modeling approach based on calculations derived from SBS demographic and denning data. We estimated that the entire SBS subpopulation produced 123 dens per year (median; 95-percent credible interval = 69, 198 dens), 66 (median; 95-percent credible interval = 35, 110 dens) of which were land-based. Most land-based dens were located between the Colville and Canning Rivers (which includes the Prudhoe Bay-Kuparuk industrial footprint), followed by the 1002 Area of the Arctic National Wildlife Refuge and the National Petroleum Reserve-Alaska.
Bromaghin, J.F., McDonald, T.L., Stirling, I., Derocher, A.E., Richardson, E.S., Rehehr, E.V., Douglas, D.C., Durner, G.M., Atwood, T. and Amstrup, S.C. 2015. Polar bear population dynamics in the southern Beaufort Sea during a period of sea ice decline. Ecological Applications 25(3):634-651. http://www.esajournals.org/doi/abs/10.1890/14-1129.1
Burns, J. J., Fay, F. H., and Shapiro, L.H. 1975. The relationships of marine mammal distributions, densities, and activities to sea ice conditions (Quarterly report for quarter ending September 30, 1975, projects #248 and 249), pp. 77-78 in Environmental Assessment of the Alaskan Continental Shelf, Principal Investiagors’ Reports. July-September 1975, Volume 1. NOAA, Environmental Research Laboratories, Boulder Colorado. pdf here.
Crockford, S.J. 2019. The Polar Bear Catastrophe That Never Happened. Global Warming Policy Foundation, London. Available in paperback and ebook formats. Crockford, S.J. 2020. State of the Polar Bear Report 2019. Global Warming Policy Foundation Report 39, London. pdf here.
Lentfer 1976. Polar bear management and research in Alaska 1974-76. Pg. 187-197 in [Anonymous]. Polar Bears: Proceedings of the 6th meeting of the Polar Bear Specialists Group IUCN/SSC, 7 December, 1976, Morges, Switzerland. Gland, Switzerland and Cambridge UK, IUCN. http://pbsg.npolar.no/en/meetings/
Regehr, E.V., Amstrup, S.C., and Stirling, I. 2006. Polar bear population status in the Southern Beaufort Sea. US Geological Survey Open-File Report 2006-1337. pdf here.
Regehr, E.V., Lunn, N.J., Amstrup, S.C., and Stirling, I. 2007. Effects of earlier sea ice breakup on survival and population size of polar bears in western Hudson Bay. Journal of Wildlife Management 71:2673-2683.
Rode, K.D., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2013. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Oral presentation by Karyn Rode, 28th Lowell Wakefield Fisheries Symposium, March 26-29. Anchorage, AK.
Rode, K.D., Regehr, E.V., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2014. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Global Change Biology 20(1):76-88. http://onlinelibrary.wiley.com/doi/10.1111/gcb.12339/abstract
Rode, K. D., R. R. Wilson, D. C. Douglas, V. Muhlenbruch, T.C. Atwood, E. V. Regehr, E.S. Richardson, N.W. Pilfold, A.E. Derocher, G.M Durner, I. Stirling, S.C. Amstrup, M. S. Martin, A.M. Pagano, and K. Simac. 2018. Spring fasting behavior in a marine apex predator provides an index of ecosystem productivity. Global Change Biology http://onlinelibrary.wiley.com/doi/10.1111/gcb.13933/full
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55 (Suppl. 1):59-76. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/42
Stirling, I. and Lunn, N.J. 1997. Environmental fluctuations in arctic marine ecosystems as reflected by variability in reproduction of polar bears and ringed seals. In Ecology of Arctic Environments, Woodin, S.J. and Marquiss, M. (eds), pg. 167-181. Blackwell Science, UK.
Stirling, I., McDonald, T.L., Richardson, E.S., Regehr, E.V., and Amstrup, S.C. 2011. Polar bear population status in the northern Beaufort Sea, Canada, 1971-2006. Ecological Applications 21:859-876. http://www.esajournals.org/doi/abs/10.1890/10-0849.1
Stirling, I., Richardson, E., Thiemann, G.W. and Derocher, A.E. 2008. Unusual predation attempts of polar bears on ringed seals in the southern Beaufort Sea: possible significance of changing spring ice conditions. Arctic 61:14-22. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/3/3
Stirling, I., Schweinsburg, R.E., Kolenasky, G.B., Juniper, I., Robertson, R.J., and Luttich, S. 1980. Proceedings of the 7th meeting of the Polar Bear Specialists Group IUCN/SSC, 30 January-1 February, 1979, Copenhagen, Denmark. Gland, Switzerland and Cambridge UK, IUCN., pg. 45-53.http://pbsg.npolar.no/en/meetings/ pdf of except here.
US Fish and Wildlife Service (USFWS) 2013. Polar Bear News 2013-14. Newsletter of the US Fish & Wildlife Service, Anchorage, Alaska. Pdf here.





You must be logged in to post a comment.