
Evolution is not just for the long-term – natural selection also goes on over short time periods. In the case of polar bears, this adaptation is almost certainly critical for its long-term survival.

Hudson Bay female with cub Wapusk National Park, Thorsten Milse, Government of Canada
Not all polar bears are identical — that is the reality that allows natural selection to operate.
I will argue that early breakup years in Western Hudson Bay weed out individual polar bears that do not have the physiological or behavioral characteristics necessary to be useful members of the population – and that this is a good thing for the entire population.

Figure 1. Slide from presentation by Julienne Stroeve, “Arctic sea ice reduction: the evidence, models, and global impacts” 22 September 2014, courtesy Ed Hawkins via Twitter. Note values prior to 1979 are vague approximations, error bars not shown (see previous post here).
Sea ice declines over the last 60 years or so have not proceeded precipitously (each year lower than the next) – there is an overall trend of less ice in September now than there was in the 1950s but with great year-to-year variability (Fig. 1, estimates to the early 1950s; and Fig. 2, satellite record, noting that 2014 will be similar to 2013).
In fact, what we have seen is that a lower-than-average ice year (at September) is generally followed by at least one higher ice cover year – and often several. The record-setting lows of 2007 and 2012 were no exception (see Fig. 2 below).

Figure 2. Average September ice extent, 1979-2013 (2014 will be similar to 2013). NSIDC.
But we know that the September minimum is not especially relevant for polar bears, since most of their feeding is done from March through June. Recall that the Chukchi Sea had very low summer/fall ice cover in 2007, yet recent research (discussed here, here, and here) showed that polar bears did very well in spite of it – they were in good condition and some females even had triplets.
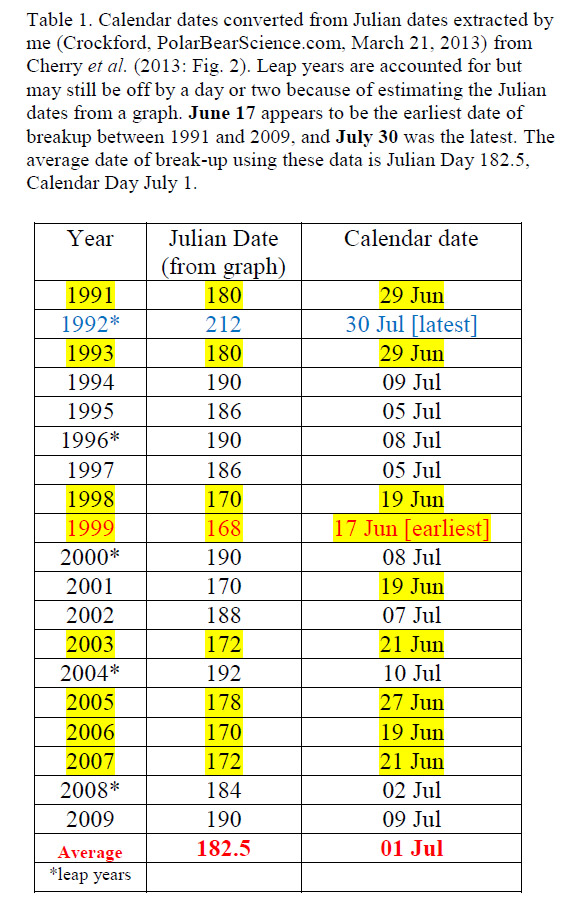
So let’s take the Western Hudson Bay polar bear region in Canada (WHB, map above from Polar Bear Specialist Group), where there has been detailed information published about ice breakup for 1991-2009 (Cherry et al. 2013). The average breakup date for WHB is July 1 (see Table 1 below), and while variable dates of freeze-up in the fall also impacts the length of the ice-free season, the pattern is the same.
The earliest breakup dates during this period occurred in 1998 and 1999 (about two weeks earlier than average) followed in 2000 by breakup that was about one week later than average. This pattern – like the one for September minimums over the entire Arctic – is repeated over the entire period.
More recently – because no one has published or announced “official” breakup dates since 2009 ̶ I’ve calculated (method described here) that 2010 was an early breakup year (similar to 1999), 2011 was a little earlier than average, and 2012 through 2014 were average or above average.
So, recent years fit the long-term pattern: there has been no consecutive “early” breakup dates (one week earlier or more) for more than two years running.
Polar bear biologist Ian Stirling and his colleague (Stirling and Parkinson 2006: 266) said this about early breakup years:
“Most polar bears can probably handle a single short ice season without a major problem. However, as the number of consecutive short ice seasons increases, the cumulative stress on the polar bears is bound to increase as well.”
While this statement is probably true, such a “worst case” scenario hasn’t happened: the bears have always had a “recovery” year (or several) to recuperate from the stress of any early breakup year or two.
Here’s the point: the variable decline-plus-recovery that has been the current reality for WHB bears is a very different situation than a steadily-declining ice cover. And because of that, the biological impact on polar bears is almost certainly going to be very different (as I suggested earlier1).
Polar bears that didn’t figure out how to hunt seals successfully over the shorter spring hunting season in early breakup years – or who didn’t have efficient fat storage metabolism due to bad genes (not their fault but that’s life) – would have starved to death.
For a few bears, death by starvation would have come at the end of the long ice-free season (November/December) but most bears destined to succumb would have died on the ice the following winter.
However, all of the bears that did survive those early breakup years – especially the females – would have included the best hunters and the bears with the best fat storage metabolism in the population.
Those were the polar bears that mated and had cubs over the next few years and contributed their superior genes to the entire population.
Every early breakup year would do the same thing – it would select out the bears with the weakest survival abilities, making the population as a whole (even if its slightly smaller) a lot more resilient to years with early breakup.
Quite simply, the fact that a few individuals die during early breakup years in Western Hudson Bay is a good thing for future polar bears, not a catastrophe.
Was this phenomenon factored into the predicted responses of polar bears to computer-calculated sea ice declines (Amstrup et al. 2008, 2011)? It doesn’t look like it to me.
Related posts:
Extirpated polar bears of St. Matthew Island spent five months on land during the summer January 21 2013
New study says polar bears could spend 2 to 4 months longer on shore than Western Hudson Bay bears do now January 29 2013
NSIDC says the sea ice minimum in 1964 was not different from 1979, 1981, or 2001 June 16 2013
Footnote 1. This is the question I asked back in July:
“Is an overall trend in breakup dates since 1979 really the same as highly variable breakup dates since 1992, as it relates to polar bear health? In other words, has the pattern we have seen in Western Hudson Bay (1-2 years when breakup was ~2 weeks early, followed by 2-3 years of average or late breakup dates, i.e., Fig. 4) really had the same effect on polar bear health over the long term as it would have if breakup dates had come relentlessly earlier and earlier over time (as implied by the statistical trend line and interpreted as “now two weeks earlier than 1980”)? Polar bear biologists imply the answer is yes – that these two phenomenon would have the same effect. But I would like to know if it’s really true or not. Are bears that have had 2-3 years to recover from a year or two of early breakup conditions really more likely to starve or lose their cubs the next time breakup comes early (because they have experienced an early breakup before), as assumed?”
References
Amstrup, S.C. 2011. Polar bears and climate change: certainties, uncertainties, and hope in a warming world. Pgs. 11-20 in R.T. Watson, T.J. Cade, M. Fuller, G. Hunt, and E. Potapov (eds.), Gyrfalcons and Ptarmigan in a Changing World, Volume 1. The Peregrine Fund, Boise, Idaho. http://dx.doi.org/10.4080/gpcw.2011.0100
Amstrup, S.C., Marcot, B.G., Douglas, D.C. 2008. A Bayesian network modeling approach to forecasting the 21st century worldwide status of polar bears. Pgs. 213-268 in Arctic Sea Ice Decline: Observations, Projections, Mechanisms, and Implications, E.T. DeWeaver, C.M. Bitz, and L.B. Tremblay (eds.). Geophysical Monograph 180. American Geophysical Union, Washington, D.C. http://onlinelibrary.wiley.com/doi/10.1029/180GM14/summary and http://alaska.usgs.gov/science/biology/polar_bears/pubs.html
Cherry, S.G., Derocher, A.E., Stirling, I., and Richardson, E.S. 2009. Fasting physiology of polar bears in relation to environmental change and breeding behavior in the Beaufort Sea. Polar Biology 32:383-391. http://link.springer.com/article/10.1007/s00300-008-0530-0#page-1
Stirling, I. and Parkinson, C.L. 2006. Possible effects of climate warming on selected populations of polar bears (Ursus maritimus) in the Canadian Arctic. Arctic 59:261-275. [open access] http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/16.





You must be logged in to post a comment.