
The trouble is, sea ice researchers and atmospheric scientists have not drawn that conclusion, despite what a new paper by Pilfold and colleagues imply. It shows just what lengths desperate IUCN Polar Bear Specialist Group (PBSG) biologists will go to in order to link the recent decline of Southern Beaufort bear numbers to global warming while ignoring similar past declines.

The authors (Pilfold, Stirling, Derocher and Richarson, 2015, in press) selected an area for their work (Fig. 1 below)1 that spans portions of two IUCN-defined polar bear subpopulations in the Eastern Beaufort Sea (Southern Beaufort and Northern Beaufort, see post here on recent boundary change made by Canada).
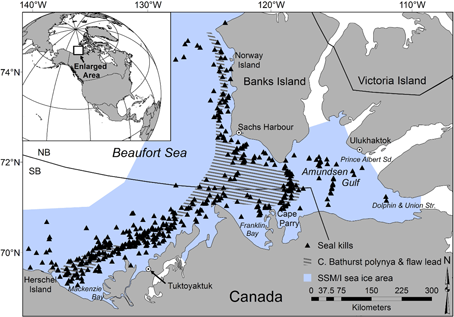
Figure 1. Area of the Pilfold et al. (2015) study in the eastern Beaufort, Canada, Fig 1 in the paper. The region encompassed part of the “Southern Beaufort” and the “North Beaufort” polar bear subpopulations.
The authors’ objective appears to have been to build a model that describes the relationship between polar bear predation on ringed seals in spring and amount of sea ice available,2
Astonishingly, although thick, highly deformed spring ice conditions are known to have been a critical factor in the repeated starvation events experienced by polar bears in the eastern Beaufort Sea on a decadal basis (going back to the 1970s and before), the authors considered only sea ice concentration (% of ice coverage) available in spring.
And although they did consider the state of the Arctic Oscillation (AO) – an Arctic-wide index not specific to the region (they said: “During negative AO winters, ice is generally thicker, resulting in colder spring temperatures and delayed melt”) – they included no references to support their implied suggestion that the condition of the AO fully explains regional conditions in the eastern Beaufort i.e., the development of thick, heavily buckled ice conditions that develop every 10 years or so in this region but not in others.
Instead, here’s what they’ve done with the AO connection – they suggest that recent extremes of the AO (e.g., extreme negative values producing cold winters and thick spring ice) could be blamed on global warming.
They cite as support for this notion a paper that dealt specifically with the North Atlantic Oscillation (NAO) and which made no claim whatsoever that the phenomenon they described could be a more general pattern for all such climatic oscillation indices. [I looked at the paper (Goodkin et al. 2008) to confirm that it makes no such claim]
This is how Pilford and colleagues link the 2004-2006 catastrophic thick spring ice conditions in the eastern Beaufort with global warming while ignoring the virtually identical conditions that occurred in 1974-1976:
“However, the effect that climatic oscillations have on sea ice may be shifting with climate change. Due to a thinning sea ice cover, a strong 2009 negative winter AO phase promoted the export of sea ice through the Chukchi Sea (Stroeve et al. 2011). Warming global temperatures may also be creating more extreme and lengthy phases in climatic oscillations (Goodkin et al. 2008). As such, future climatic oscillation patterns may result in unexpected consequences for polar bear foraging ecology that do not reflect past responses.” [my bold]
They had this to say about the implications of their model results — notice they don’t deny that bear survival was strongly correlated with thick deformed spring ice and a scarcity of ringed seal pups in 2004-2006, they just don’t mention the fact that the same thing happened in the 1970s (Stirling and Lunn 1997; Stirling 2002):
“Ice in the Beaufort Sea in the mid-2000s had extensive rubble zones, which may have made hunting more difficult (Stirling et al. 2008). Further study on differentiating between the abundance and the accessibility of seals as it relates to climate and predation is warranted. Because polar bears may acquire as much as two-thirds of their energetic intake in spring (Stirling and Øritsland 1995), a significant reduction in kill biomass could have population level consequences. Spring is the mating season for polar bears, and successfully mated females can avoid implantation or abort fetuses before denning if fat stores are inadequate (Derocher et al. 1992). Additionally, Bromaghin et al. (2014) reported lower survival rates between 2004 – 2006 for the southern Beaufort subpopulation and suggested measures of ice-availability were insufficient in explaining the decline. We suggest that poor hunting conditions in spring 2005 – 2006 could have negatively affected polar bear survival.”[my bold]
They concluded:
“Results suggest documented changes in polar bear fasting rates in the Beaufort Sea, Canada are likely due to a complex set of abiotic and biotic factors including potential underlying prey population dynamics.”
Pilfold and colleagues’ attempt to model a link between the 2004-2006 polar bear decline in the Southern Beaufort to global warming, while ignoring the virtually identical 1974-1976 event that occurred before much warming had occurred — similar to what Stirling and colleagues attempted to do in a previous (2008) paper — is scientifically unsound and unsupported. So much for the value of peer review.
If sea ice experts and atmospheric scientists had been able to link the development of thick, buckled spring ice conditions in the Eastern Beaufort to global warming — and then to starving polar bears — I am absolutely certain we’d have heard about it by now. The fact that they have not indicates that these polar bear biologists are grasping at straws to connect recent polar bear population declines in the Southern Beaufort to global warming while ignoring identical prior events.
Could it be that the publication of this paper had something to do with the fact that PBSG biologists need many bits of “evidence” to include in the model they need for their 2015 IUCN Red List assessment (due in June) so that they can continue to predict a dire future for polar bears because of global warming? [note that all co-authors of this paper are IUCN PBSG members].
Related posts:
Great polar bear red herring in the Southern Beaufort Posted July 4, 2013
Where were the appeals to feed starving polar bears in 1974? February 21, 2013
Why is it that every decade, Eastern Beaufort sea ice gets really thick? June 28, 2013
Polar bear researchers knew S. Beaufort population continued to increase up to 2012 November 19, 2014
Cooling the polar bear spin July 26, 2012
IUCN Polar Bear Specialist Group out-lived its usefulness 20 years ago January 1, 2015
Amstrup knows his polar bear predictions are flawed – but continues to promote them November 29, 2014
Footnote 1: Here’s how the authors describe the polar bear data they collected:
“The study area comprised the eastern Beaufort Sea, including Amundsen Gulf, within 150 km of the Canadian coast (Fig. 1). Helicopter flights originated from Tuktoyaktuk, Sachs Harbour, Ulukhaktok (Holman), Cape Parry and Norway Island, Northwest Territories. Observations of seals killed by polar bears were collected over 16 springs between early-April and late-May (range 3 April 28 May) in 1985–1987, 1992–1994, 2000, and 2003–2011. The study area is composed mainly of annual fast ice along the coast and pack ice farther off shore, separated by the Cape Bathurst polynya and related flaw lead system. The flaw lead system is biologically productive with high seal abundance (Stirling et al. 1982).”
Footnote 2: The objective of the paper is described this way:
“At a larger temporal scale, the physical structure of sea ice in spring is reflective of climatic conditions over the winter (Rigor et al. 2002), and may influence predation events (Stirling et al. 2008). Finally, to examine the effect of climate change on predation, we integrated the results of our study with findings on fasting rates in polar bears. Cherry et al. (2009) reported that the proportion of polar bears fasting in the spring in the Beaufort Sea had increased from 10% in 1985 – 1986 to 25% in 2005 – 2006. We applied the conditions observed in these periods to the top predation model to examine if the expected kill rate from the model could explain the observed polar bear fasting rates.” [my bold]
The paper:
Pilfold, N. W., Derocher, A. E., Stirling, I. and Richardson, E. 2015 in press. Multi-temporal factors influence predation for polar bears in a changing climate. Oikos. doi: 10.1111/oik.02000 Article first published online: 6 FEB 2015. http://onlinelibrary.wiley.com/doi/10.1111/oik.02000/abstract
Abstract [my bold]
Predation is an ecological interaction influenced by abiotic and biotic factors acting on multiple temporal scales, yet multi-temporal comparisons are rare in empirical studies. For polar bears Ursus maritimus, the physical configuration of the habitat and conditions in which seals are hunted may change on intra- and inter-seasonal scales. Additionally, while the effects of climate change on polar bears have focused on linking reductions in sea ice to body condition and survival, the potential changes to on-ice hunting conditions have not been examined. Employing observational counts of seals killed by polar bears between early-April and late-May 1985–2011 (n = 650), we modelled the likelihood of predation events in the Beaufort Sea, Canada at multi-temporal scales. We used the top model to estimate the expected kill rate of seals in the springs of 1985–1986 and 2005–2006 and integrated the result with fasting rates derived from physiological markers in blood samples. A log-likelihood ratio test suggested a multi-temporal approach fit the seal kill data better than any single scale alone. Predation events were influenced by ringed seal Pusa hispida reproduction and haul-out behaviour, regional sea ice concentration and the phase of climatic indices. The expected kill rate from the top predation model and the estimated mean biomass of seal kills were significant predictors of polar bear fasting rates. Results suggest that 50% less seal biomass was killed in 2005–2006 than in 1985–1986, which correlates with a significant increase in the frequency of polar bears in a fasting state. We propose that the documented changes in polar bear fasting rates between 1985–1986 and 2005–2006 are due to a complex set of abiotic and biotic factors including underlying prey dynamics, rather than a single-scale environmental correlation.
Other References
Cherry, S.G., Derocher, A.E., Stirling, I., and Richardson, E.S. 2009. Fasting physiology of polar bears in relation to environmental change and breeding behavior in the Beaufort Sea. Polar Biology 32:383-391. http://link.springer.com/article/10.1007/s00300-008-0530-0#page-1
Goodkin, N.F., Hughen, K.A., Doney, S.C. and Curry, W.B. 2008. Increased multidecadal variability of the North Atlantic Oscillation since 1781. Nature Geoscience 1:844 – 848. http://www.nature.com/ngeo/journal/v1/n12/abs/ngeo352.html
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55 (Suppl. 1):59-76. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/42
Stirling, I. and Derocher, A.E. 2013. Effects of climate warming on polar bears: a review of the evidence. Global Change Biology 18 (9): 2694-2706 doi:10.1111/j.1365-2486.2012.02753.x http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2486.2012.02753.x/abstract
Stirling, I. and Lunn, N.J. 1997. Environmental fluctuations in arctic marine ecosystems as reflected by variability in reproduction of polar bears and ringed seals. In Ecology of Arctic Environments, Woodin, S.J. and Marquiss, M. (eds), pg. 167-181. Blackwell Science, UK.
Stirling, I., Richardson, E., Thiemann, G.W. and Derocher, A.E. 2008. Unusual predation attempts of polar bears on ringed seals in the southern Beaufort Sea: possible significance of changing spring ice conditions. Arctic 61:14-22. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/3/3





You must be logged in to post a comment.