
![]() We know that thick-ice springs occurred in 1974, 1975, 1986, 1992, 2004, and 2005 in the former ‘Eastern Beaufort’ – now the southern portion of the ‘Northern Beaufort’ and the eastern portion of the ‘Southern Beaufort.’ We know that these severe spring ice conditions negatively impacted both polar bears and ringed seals in this region every decade since the 1960s because the effects have been documented by numerous studies conducted in April through May for polar bears (Amstrup et al. 2006; Cherry et al. 2009; Pilfold et al. 2012; Stirling 2002; Stirling and Lunn 1997; Stirling et al. 1980; Stirling et al. 1993; Stirling et al. 2008) and in June and July for ringed seals (Harwood et al. 2012; Smith 1987), see previous posts here, here, and here.
We know that thick-ice springs occurred in 1974, 1975, 1986, 1992, 2004, and 2005 in the former ‘Eastern Beaufort’ – now the southern portion of the ‘Northern Beaufort’ and the eastern portion of the ‘Southern Beaufort.’ We know that these severe spring ice conditions negatively impacted both polar bears and ringed seals in this region every decade since the 1960s because the effects have been documented by numerous studies conducted in April through May for polar bears (Amstrup et al. 2006; Cherry et al. 2009; Pilfold et al. 2012; Stirling 2002; Stirling and Lunn 1997; Stirling et al. 1980; Stirling et al. 1993; Stirling et al. 2008) and in June and July for ringed seals (Harwood et al. 2012; Smith 1987), see previous posts here, here, and here.
For example, even though Ian Stirling and colleagues argued in their 2008 paper that the thick spring ice conditions in 2004, 2005 and 2006 (but not those in previous decades) were caused by storms initiated or intensified by greater amounts of open water in previous summers, they did not deny that the thick-ice springs occurred. They stated quite clearly that:
“The 1960s, 1970s, and 1980s each experienced a two- to three-year decline in seal productivity in the eastern Beaufort Sea and Amundsen Gulf, associated with heavy ice conditions, around mid-decade. Each was followed by a decline in polar bear reproduction and condition, after which both seal and bear populations recovered (Smith, 1987; Harwood et al., 2000; Stirling, 2002). The beginning of each of those three periods was associated with heavy ice conditions through the winter before the reproductive decline of the seals, followed by a late spring breakup.” [my bold]
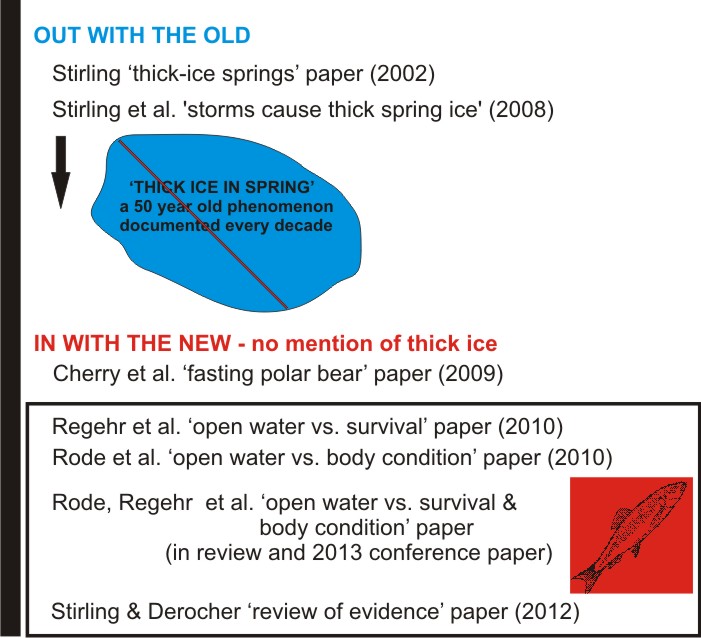
So, I have to say, I was shocked but not surprised to find that in the more recent scientific literature, the phenomenon of thick-ice springs every decade in Southern and Northern Beaufort has been deliberately ‘disappeared.’
Not surprised because I suspected it had happened — this issue was a feature of the Stirling and Derocher (2012) paper from late last year which was the topic of my very first blog post, “Cooling the polar bear spin.”
However, I think it is important to document how the transmogrification of sea ice effects on polar bears was managed in the scientific literature so that everyone can see exactly what has been done. In a truly astonishing move for what is supposed to be a field of science, thick-ice springs have been effectively replaced by an open-water red herring as the scourge of Southern Beaufort polar bears.
Shortly after the Stirling et al. paper (quoted from above) was published in 2008, Alaskan polar bear researchers abandoned the implausible ‘storms cause thick spring ice’ proposal that it included, probably because it accepted the premise that thick-ice springs were the ultimate cause of polar bear woes in the Beaufort Sea. Not only was the ‘storms cause thick spring ice’ premise a weak argument, it also didn’t fit the big-picture story that was being developed at the time: that declines in summer sea extent are the biggest threat to polar bears everywhere (see previous post here).
Instead, the declines in polar bear body condition and cub/yearling survival of polar bears that occurred in the mid-2000s are now being blamed on the modest decline in sea ice extent in the late summer that have occurred over the last 20 years or so. The starving bears of the 1970s are no longer a concern – it’s the starving bears of the 2000s that matter now.
The year after the Stirling et al. ‘storms cause thick spring ice’ (2008) paper was published, student lead author Seth Cherry and colleagues (Derocher, Stirling and Richardson) published a paper on polar bear fasting in the Southern Beaufort, which I’ll call a “transition” paper. It forms a bridge between the weak ‘storms cause thick spring ice’ premise and the preferred red herring storyline, ‘more open water causes polar bear declines.’
Cherry et al. examined indicators of fasting in blood samples taken from bears in the Eastern Beaufort [there is the ‘Eastern Beaufort’ again as a study area but not a formal subpopulation] during April and May of 1985/1986 and 2005/2006. They found:
“the proportions of polar bears fasting were 9.6% in 1985, 10.5% in 1986, 21.4% in 2005, and 29.3% in 2006….The increased number of polar bears in a physiological fasting state from all sex, age, and reproductive classes in 2005 and 2006 corresponded with broad scale changes in Arctic sea ice composition, which may have affected prey availability.” [my bold]
Feigning ignorance of past studies (even though Ian Stirling was a co-author), the ‘fasting’ paper concluded that:
“the recent increase in bears fasting during April and May suggests that there has been a decrease in prey availability at that time of year. …the precise means through which prey have apparently become less available to polar bears in the Beaufort Sea remains unknown.” [my bold]
Cherry et al. went on to infer, without actually coming out and saying so, that the likely cause of the increased fasting in the spring of 2005 and the spring of 2006 was reduced ice in summer and more open water.
A useful bridge has to lead somewhere and the student-led fasting paper (Cherry et al.) was followed in 2010 by one written by a much more experienced research team. Led by by Eric Regehr (with Ian Stirling again one of the co-authors) “evaluated the effects of sea ice conditions on vital rates (survival and breeding probabilities) for polar bears in the southern Beaufort Sea,” using models. It was based on only five years of data (collected from 2001 to 2006) on polar bears and number of ice-free days each year. They concluded:
“In the most supported models, polar bear survival declined with an increasing number of days per year that waters over the continental shelf were ice free. In 2001–2003, the ice-free period was relatively short (mean 101 days) and adult female survival was high (0·96–0·99, depending on reproductive state). In 2004 and 2005, the ice-free period was longer (mean 135 days) and adult female survival was low (0·73–0·79, depending on reproductive state). Breeding rates and cub litter survival also declined with increasing duration of the ice-free period. Confidence intervals on vital rate estimates were wide.” [my bold]
Unbelievable. No mention at all of the 50-year history of thick-ice springs in this region, replaced by a handy ‘correlation‘ with open water. [See also previous post here for a critique of the mark-recapture methods and models used in this study]
Because Regehr et al. did not consider whether thick ice in spring had an effect on polar bear survival in their 2010 paper, they found a correlation with increased open water a few months later in the year – hardly surprising. Hardly surprising but also, hardly scientific.
This correlation is a blatant red herring: bears in very poor condition in May (especially cubs and independent subadults) because of thick spring ice were almost certainly still in relatively poor condition three months later. The correlation is valid, the implied causation is not.
A companion paper (Rode et al. 2010) on polar bear body condition and growth in the Southern Beaufort does something similar as Regehr et al.’ s ‘open water vs. survival’ paper – both cite the other as corroborative evidence, and both reference the Cherry et al. ‘fasting’ paper.
The Rode et al. 2010 ‘open water vs. body condition’ paper used data from a longer period (1982 to 2006) and looked at changes over time in body condition (relative fatness) and growth patterns of polar bears in relation to changes in the availability of ice areas (in km2) “most frequently used by polar bears.” The study did not address sea ice conditions per se, and as for the Regehr et al. 2010 ‘open water vs. survival’ study, nowhere in the paper was the phenomenon of thick-ice springs even mentioned.
Finally, as confirmation that the bridge to the open water red herring has been successfully built, we now have an “in review” paper co-authored by Rode, Regehr and others waiting in the wings (summarized by Rode et al. 2013, a conference presentation), entitled “Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations.” I’ll have more to say about it later.
This ‘in review’ paper merges the Regehr et al. 2010 ‘open water vs. survival’ and Rode et al. 2010 ‘open water vs. body condition’ conclusions as fact and develops the entire storyline: recent declines in body condition and survival of polar bears in the Southern Beaufort are correlated with more open water and declining sea ice caused by global warming – with a very strong implication that ‘correlated’ means ‘caused by.’
Voilà — a perfectly prepared red herring (summarized below).
Finally, we have the Stirling and Derocher ‘review of evidence’ paper [summarizing the “documented” effects of global warming on polar bears, now in print], which neatly foresaw this transmogrification and integrated it into their big-picture story.
The scientific paper trail for the red herring is now complete.
And to avoid any inconvenient data related to the thick-ice springs phenomenon arising in future, the focus of research effort in the Southern Beaufort has been shifted from spring to late summer/early fall (Fish & Wildlife Service 2009; Polar Bear News 2010; Regehr et al. 2010; Rode and Regehr 2010).
References
Amstrup, S.C., Stirling, I., Smith, T.S., Perham, C. and Thiemann, B.W. 2006. Recent observations of intraspecific predation and cannibalism among polar bears in the Southern Beaufort Sea. Polar Biology 29:997–1002. Pdf here.
Cherry, S.G., Derocher, A.E., Stirling, I., and Richardson, E.S. 2009. Fasting physiology of polar bears in relation to environmental change and breeding behavior in the Beaufort Sea. Polar Biology 32:383-391. http://link.springer.com/article/10.1007/s00300-008-0530-0#page-1
Fish and Wildlife Service 2009. Ongoing polar bear research in Alaska. US Fish & Wildlife, Anchorage, AK. Pdf here.
Harwood, L.A., Smith, T.G., Melling, H., Alikamik, J. and Kingsley, M.C.S. 2012. Ringed seals and sea ice in Canada’s western Arctic: harvest-based monitoring 1992-2011. Arctic 65:377-390. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/4236
Pilfold, N.W., Derocher, A.E., Stirling, I., Richardson, E., and Andriashek, D. 2012. Age and sex composition of seals killed by polar bears in the eastern Beaufort Sea. PLoS ONE 7:e41429. Doi:10.1371/journal.pone.0041429.
Polar Bear News. 2010. Polar bear newsletter of the US Fish & Wildlife Service, Anchorage, Alaska. Pdf here.
Regehr, E.V., Hunter, C.M., Caswell, H., Amstrup, S.C., and Stirling, I. 2010. Survival and breeding of polar bears in the southern Beaufort Sea in relation to sea ice. Journal of Animal Ecology 79:117–127. http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2656.2009.01603.x/full
Rode, K.D., Amstrup, S.C., and Regehr, E.V. 2010. Reduced body size and cub recruitment in polar bears associated with sea ice decline. Ecological Applications 20:768-782. http://www.esajournals.org/doi/abs/10.1890/08-1036.1
Rode, K.D., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2013. Comparison in polar bear response to sea ice loss in the Chukchi and southern Beaufort Seas. Oral presentation at the 28th Lowell Wakefield Fisheries Symposium, March 26-29. Anchorage, AK. [more on this later, it was a review of Rode et al., in review, below]
Rode, K.D., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. in review [journal not stated]. “Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations.” [cited in Rode et al. 2013 above]
Rode, K. and Regehr, E.V. 2010. Polar bear research in the Chukchi and Bering Seas: A synopsis of 2010 field work. Unpublished report to the US Fish and Wildlife Service, Department of the Interior, Anchorage. Pdf here. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/42
Smith, T.G. 1987. The Ringed Seal, Phoca hispida, of the Canadian Western Arctic. Canadian Bulletin of Fisheries and Aquatic Sciences 216. Department of Fisheries and Oceans, Ottawa. Google Books link http://tinyurl.com/ppqrf6k
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55 (Suppl. 1):59-76. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/42
Stirling, I. and Derocher, A.E. 2012. Effects of climate warming on polar bears: a review of the evidence. Global Change Biology 18 (9): 2694-2706 doi:10.1111/j.1365-2486.2012.02753.x http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2486.2012.02753.x/abstract
Stirling, I. and Lunn, N.J. 1997. Environmental fluctuations in arctic marine ecosystems as reflected by variability in reproduction of polar bears and ringed seals. In Ecology of Arctic Environments, Woodin, S.J. and Marquiss, M. (eds), pg. 167-181. Blackwell Science, UK.
Stirling, I., Andriashek, D., and Calvert, W. 1993. Habitat preferences of polar bears in the western Canadian Arctic in late winter and spring. Polar Record 29:13-24. http://tinyurl.com/qxt33wj
Stirling, I., McDonald, T.L., Richardson, E.S., Regehr, E.V., and Amstrup, S.C. 2011. Polar bear population status in the northern Beaufort Sea, Canada, 1971-2006. Ecological Applications 21:859-876. http://www.esajournals.org/doi/abs/10.1890/10-0849.1
Stirling, I., Richardson, E., Thiemann, G.W. and Derocher, A.E. 2008. Unusual predation attempts of polar bears on ringed seals in the southern Beaufort Sea: possible significance of changing spring ice conditions. Arctic 61:14-22. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/3/3
Stirling, I., Schweinsburg, R.E., Kolenasky, G.B., Juniper, I., Robertson, R.J., and Luttich, S. 1980. Proceedings of the 7th meeting of the Polar Bear Specialists Group IUCN/SSC, 30 January-1 February, 1979, Copenhagen, Denmark. Gland, Switzerland and Cambridge UK, IUCN., pg. 45-53.http://pbsg.npolar.no/en/meetings/ pdf of except here.





You must be logged in to post a comment.