
As well as summarizing sea ice changes, NOAA’s 2020 Arctic Report Card features two reports that document the biggest advantage of much less summer sea ice than there was before 2003: increased primary productivity. Being at the top of the Arctic food chain, polar bears have been beneficiaries of this phenomenon because the Arctic marine mammals they depend on for food – seals, walrus and bowhead whales – have been thriving despite less ice in summer.
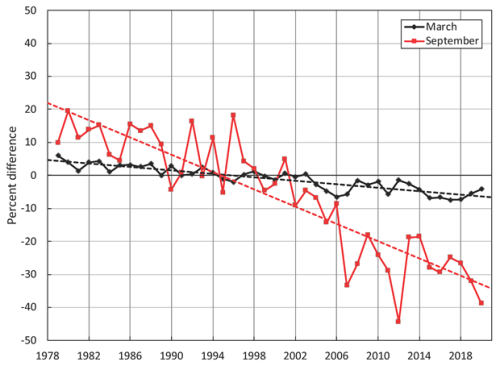
In the sea ice chapter (Perovich et al. 2020), my favourite of all the figures published is the graph of September vs. March sea ice (above). As you can see, March ice extent has been virtually flat (no declining trend) since 2004. And as the graph below shows, September extent has been without a trend since 2007, as NSIDC ice expert Walt Meier demonstrated last year (see below): it doesn’t take much imagination to see that the value for 2020 from the graph above (the second-lowest after 2012) hasn’t changed the flat-trend line.

The chapter also included this comparison of March vs. September ice charts:
 However, the real eye-opener in the report is the admission that much less summer ice has benefitted the entire Arctic food change because of increased primary productivity.
However, the real eye-opener in the report is the admission that much less summer ice has benefitted the entire Arctic food change because of increased primary productivity.
Benefits of less summar ice
One of the highlights emphasized in the report is this gem:
During July and August 2020, regional ocean primary productivity in the Laptev Sea was ~2 times higher for July and ~6 times higher for August compared to their respective monthly averages.
It’s nice to see an acknowledgement that the longer ice-free seasons we’ve experienced since 2007 have an up-side. In fact, less summer ice has been a net benefit to most animals in the Arctic and peripheral seas because less ice and more sunlight in most areas increases ‘primary productivity’. Primary productivity refers to phytoplankton, those single-celled plants that are the basis for life in the ocean because they turn sunlight into stored energy: sunlight is their food. Longer ice-free seasons – featuring less ice and more sunlight – provide the conditions phytoplankton need to grow exponentially, producing ‘blooms’ that can be seen by satellites. One such bloom is shown below, in the Barents Sea on 26 July 2020 (NOAA photo).

In the table presented by one set of NOAA authors (Frey et al. 2020) below, primary productivity has been up since 2003, not including data from 2020, in virtually all regions of the Arctic:

Similar results are discussed in a government report from Canada (Coupel et al. 2019), which summarizes reports of recent phytoplankton increases across the Arctic. I’ve revised a graphic I posted earlier this year to show how this works (below). More primary productivity due to a longer ice-free period benefits the entire Arctic food chain in which polar bears hold top spot: fatter seals because of more food mean fatter polar bears with improved survival.

Abundant phytoplankton → more food for single-celled animals (zooplankton), fish, and bottom-dwelling invertebrates like clams → prolific reproduction of krill, fish and clams (population increases); more fish, in turn → fatter ringed and bearded seals, who feed primarily in the ice-free season, and thus fat female seals → more fat pups the following spring (Crawford et al. 2015) for polar bears when they need it most. Unless other factors come into play that reduce prey availability, like too much ice, or snow over ice in spring (Crockford 2015, 2017), polar bears will tend to be fatter, healthier, and reproduce more successfully, resulting in at least stable, if not growing population numbers – such as we’ve seen in the Barents and Chukchi Seas, and in the Gulf of Boothia, since 2005 (Aars 2018; Dyck et al. 2020; Lippold et al. 2019; Regehr et al. 2018; Rode et al. 2014, 2018).
As well as ringed seals, bearded seals and polar bears, bowhead whales (shown below) have also benefitted from this increased primary productivity, as explained in another chapter in the Arctic Report Card by John George and colleages.

These authors (George et al. 2020) stated in their chapter about bowhead whales (see range map from their paper below):
The population size of bowheads in the Pacific Arctic has increased in the past 30 years likely due to increases in ocean primary production as well as the northward transport of the zooplankton on which they feed.

In addition, I suspect that the large recent population size and health of Pacific walrus are indicators they are another species that has been the beneficiary of less summer sea ice (Crockford 2014a,b). Walrus feed on bottow-dwelling invertebrate creatures whose population sizes would be boosted by more abundant plankton, allowing more walrus to forage without running out of food, as they have been known to do in the past during so-called ‘boom and bust’ population cycles (Fischbach et al. 2016; Lowry 1985; MacCracken et al. 2017).
References
Aars, J. 2018. Population changes in polar bears: protected, but quickly losing habitat. Fram Forum Newsletter 2018. Fram Centre, Tromso. Download pdf here (32 mb).
Coupel, P., Michel, C. and Devred, E. 2019. Case study: The Ocean in Bloom. In State of Canada’s Arctic Seas, Niemi, A., Ferguson, S., Hedges, K., Melling, H., Michel, C., et al. 2019. Canadian Technical Report Fisheries and Aquatic Sciences 3344.
Crawford, J.A., Quakenbush, L.T. and Citta, J.J. 2015. A comparison of ringed and bearded seal diet, condition and productivity between historical (1975–1984) and recent (2003–2012) periods in the Alaskan Bering and Chukchi seas. Progress in Oceanography 136:133-150.
Crockford, S. J. 2014a. On the beach: walrus haulouts are nothing new. Global Warming Policy Foundation Briefing Paper 11. Pdf here.
Crockford, S. J. 2014b.The walrus fuss: walrus haulouts are nothing new http://www.thegwpf.org/gwpftv/?tubepress_item=cwaAwsS2OOY&tubepress_page=2
Crockford, S.J. 2015. The Arctic Fallacy: Sea Ice Stability and the Polar Bear. Global Warming Policy Foundation Briefing Paper 16. London. Available at http://www.thegwpf.org/susan-crockford-the-arctic-fallacy-2/
Crockford, S.J. 2017. Testing the hypothesis that routine sea ice coverage of 3-5 mkm2 results in a greater than 30% decline in population size of polar bears (Ursus maritimus). PeerJ Preprints 19 January 2017. Doi: 10.7287/peerj.preprints.2737v1 Open access. https://peerj.com/preprints/2737/
Dyck, M., Regehr, E.V. and Ware, J.V. 2020. Assessment of Abundance for the Gulf of Boothia Polar Bear Subpopulation Using Genetic Mark-Recapture. Final Report, Government of Nunavut, Department of Environment, Iglulik. 12 June 2020. Pdf here.
Fischbach, A.S., Kochnev, A.A., Garlich-Miller, J.L., and Jay, C.V. 2016. Pacific walrus coastal haulout database, 1852–2016—Background report: U.S. Geological Survey Open-File Report 2016–1108. http://dx.doi.org/10.3133/ofr20161108. The online database is found here.
Frey, K.E., Comiso, J.C., Cooper, L.W., Grebmeier, J.M. and Stock, L.V. 2020. Arctic Ocean primiary productivity: the response of marine algae to climate warming and sea ice decline. 2020 Arctic Report Card. NOAA. DOI: 10.25923/vtdn-2198 https://arctic.noaa.gov/Report-Card/Report-Card-2020/ArtMID/7975/ArticleID/900/Arctic-Ocean-Primary-Productivity-The-Response-of-Marine-Algae-to-Climate-Warming-and-Sea-Ice-Decline
George, J.C., Moore, S.E. and Thewissen, J.G.M. 2020. Bowhead whales: recent insights into their biology, status, and resilience. 2020 Arctic Report Card, NOAA. DOI: 10.25923/cppm-n265 https://arctic.noaa.gov/Report-Card/Report-Card-2020/ArtMID/7975/ArticleID/905/Bowhead-Whales-Recent-Insights-into-Their-Biology-Status-and-Resilience
Lippold, A., Bourgeon, S., Aars, J., Andersen, M., Polder, A., Lyche, J.L., Bytingsvik, J., Jenssen, B.M., Derocher, A.E., Welker, J.M. and Routti, H. 2019. Temporal trends of persistent organic pollutants in Barents Sea polar bears (Ursus maritimus) in relation to changes in feeding habits and body condition. Environmental Science and Technology 53(2):984-995.
Lowry, L. 1985. “Pacific Walrus – Boom or Bust?” Alaska Fish & Game Magazine July/August: 2-5. pdf here.
MacCracken, J.G., Beatty, W.S., Garlich-Miller, J.L., Kissling, M.L and Snyder, J.A. 2017. Final Species Status Assessment for the Pacific Walrus (Odobenus rosmarus divergens), May 2017 (Version 1.0). US Fish & Wildlife Service, Anchorage, AK. Pdf here (8.6 mb).
Perovich, D., Meier, W., Tschudi, M., Hendricks, S., Petty, A.A., Divine, D., Farrell, S., Gerland, S., Haas, C., Kaleschke, L., Pavlova, O., Ricker, R., Tian-Kunze, X., Webster, M. and Wood, K. 2020. Sea ice. 2020 Arctic Report Card, NOAA. https://arctic.noaa.gov/Report-Card/Report-Card-2020/ArtMID/7975/ArticleID/891/Sea-Ice Pdf of entire Arctic Report Card here (12mb).
Regehr, E.V., Hostetter, N.J., Wilson, R.R., Rode, K.D., St. Martin, M., Converse, S.J. 2018. Integrated population modeling provides the first empirical estimates of vital rates and abundance for polar bears in the Chukchi Sea. Scientific Reports 8 (1) DOI: 10.1038/s41598-018-34824-7 https://www.nature.com/articles/s41598-018-34824-7
Rode, K.D., Regehr, E.V., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2014. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Global Change Biology 20(1):76-88. http://onlinelibrary.wiley.com/doi/10.1111/gcb.12339/abstract
Rode, K. D., R. R. Wilson, D. C. Douglas, V. Muhlenbruch, T.C. Atwood, E. V. Regehr, E.S. Richardson, N.W. Pilfold, A.E. Derocher, G.M Durner, I. Stirling, S.C. Amstrup, M. S. Martin, A.M. Pagano, and K. Simac. 2018. Spring fasting behavior in a marine apex predator provides an index of ecosystem productivity. Global Change Biology http://onlinelibrary.wiley.com/doi/10.1111/gcb.13933/full





You must be logged in to post a comment.