
I have a new paper out that explains a fundamental problem with polar bear conservation.

I’m convinced that a flawed and out-dated ecological concept — that sea ice, under natural conditions, provides a stable, predictable habitat — is what has allowed the present doom and gloom attitude of most polar bear specialists to develop.
Sea ice changes, of course, from season to season. However, the concept that sea ice is a stable habitat assumes that these seasonal changes are predictable and virtually the same from one year to the next – at least, similar enough that the differences are not responsible for causing marked declines in population size.
The assumption is that under natural, stable conditions populations of Arctic animals will either stay the same over time or increase. Biologists were taught at university that sea ice should be a stable habitat and as a result, they’ve glossed over evidence they collected to the contrary. [see recent posts here and here, for example]
Negative effects on populations of short-term natural variations in spring sea ice or spring snow cover on sea ice have been entirely ignored in modeled predictions of future conditions. The focus has been on summer ice extent.
I have summarized this evidence in a fully referenced, peer-reviewed essay that explores how the acceptance of this fallacy (“sea ice is a stable habitat”) has so skewed the conservation biology of polar bears that to outsiders it may look like a scientific integrity issue.
The summary and the essay are below (with embedded links and references). The Global Warming Policy Foundation (GWPF) has published the essay in their “Briefing Paper” series (#16, The Arctic Fallacy: Sea Ice Stability and the Polar Bear), which includes a must-read foreword by Dr. Matthew Cronin, Professor of Animal Genetics at the University of Alaska Fairbanks. Press release here, pdf here.
I think you’ll find it timely and thought-provoking.
The Arctic Fallacy: Sea Ice Stability and the Polar Bear
Summary
Since the late 1960s, Arctic marine mammal conservation has been based on the assumption that sea ice provides a stable, predictable environment for polar bears and Arctic seals: today, it underpins their ‘threatened with extinction’ status. A stable environment, the oversimplified K-selection theory goes, should support populations at relatively high levels over time, without marked variation in size due to habitat change.
This idealized concept was strongly promoted by the most popular university-level ecology textbooks of the 1970s and was embraced by early polar bear biologists, who began their careers at a time when polar bear were truly threatened with extinction by overhunting.
Observations since then, however, have shown the assumption of sea ice as a stable habitat over short time scales is false. Spring sea ice thickness has been naturally variable over time scales of a few years to decades in the Beaufort Sea, East Greenland, and Hudson Bay; spring ice extent has been naturally variable in the Barents Sea for centuries and spring snow depth on sea ice is known to vary over short periods.
Marked declines in polar bear and ringed seal survival in response to thick spring sea ice and reduced snow depth have been documented. These two variables are closely tied because spring (April — June) is the period of on-ice birth and nursing for ice-dependent seals and is also when polar bears consume two-thirds of their annual prey.
Apparently expecting stable or increasing populations, despite their own evidence to the contrary, Arctic biologists now surprisingly attribute virtually every downturn in population size of Arctic species to declines in summer sea ice blamed on human use of fossil fuels.
Shifting the blaming for the devastation caused by thick spring ice onto recent summer ice declines, biologists portray summer ice changes as manifestations of unprecedented, human-caused habitat instability.
Regardless of such willful blindness to the facts, the assumption that Arctic sea ice is a naturally stable habitat over short time frames is a biological fallacy. Predictive population models based on this myth are flawed, their results illusory. Yet, the International Union for the Conservation of Nature (IUCN) and the US government have, for the first time, accepted modeled (future) population declines of Arctic species based on modeled (future) summer sea ice changes as valid threats to their survival, all built upon this fallacy.
Given what we now know about the animals and their naturally changing habitat, it is time to concede that data do not support predictions that polar bears, walrus, and Arctic seals are threatened with extinction due to habitat instability.
Section 1. Introduction
Research used to support polar bears’ status as ‘threatened’ under the American Endangered Species Act (ESA), and ‘vulnerable’ by the International Union for the Conservation of Nature (IUCN)(US Fish and Wildlife Service 2008; Schliebe et al. 2008) is based on the concept that Arctic sea ice is such a stable habitat that no profound fluctuations in population size ever take place naturally as a result of seasonal changes in sea ice. In other words, unlike terrestrial habitats such as grasslands or deserts, where periodic heavy winter snowfall or summer drought may cause animal populations to crash (e.g. Martinka 1967; Anthony 1976), the inherent stability of sea ice in all seasons is assumed to buffer polar bears and Arctic seals from such natural population fluctuations. How did such a bizarre idea – that sea ice habitat never varies naturally, even over short time periods – come to underpin a declaration that a species was ‘threatened’ (Adler 2008; Akçakaya et al. 2006)?
In the 1960s, some biologists who studied animal populations looked for an overarching pattern of interaction between animals and their habitats. One ecological feature of particular interest was the ability of the habitat to support populations of animals over time, the habitat’s so-called ‘carrying-capacity’, referred to as ‘K’.
By the early 1970s, when research on Arctic seals, walrus and polar bears began in earnest, a popular yet short-lived paradigm had emerged. This divided all species into so-called ‘r-selected’ or ‘K-selected’ groups.
The category of r-selected animals includes those with ‘many offspring, short gestation, less parental care, and a short time until sexual maturity’ and which live in unstable or unpredictable habitats. Examples include bacteria or mice. K-selected species are those with ‘few offspring, long gestation, long parental care, and a long period until sexual maturity’ and which live in stable, predictable habitats. Examples include whales and humans. This concept was strongly promoted by the most popular university-level ecology textbooks of the 1970s (Reznick et al. 2002: 1511).
Polar bears and walrus were then, and still are today, considered to be classic K-selected species8 by the specialists who study them (e.g., Vongraven et al. 2012: 2, 17; Laidre et al.2008), as shown by this statement made more than a decade ago by Derocher and colleagues (Derocher et al. 2004:163):
Polar bears (Ursus maritimus) are a classic K-selected species having delayed maturation, small litter sizes, and high adult survival rates.
While unbridled human predation on walrus and polar bears in the 19th and early 20th centuries temporarily made the notion of a ‘balanced’ Arctic ecosystem invalid, Arctic biologists believed that recovered populations should reach and then maintain a constant level in their stable habitat: populations would not decline due to naturally-occurring changes in their habitat because such environmental changes would not happen, even over the short term.
For carnivores such as polar bears, the theory that stable habitats support stable population numbers over short timescales also assumed the presence of a relatively constant number of animals to feed upon – in other words, a stable prey base of Arctic seals, primarily ringed and bearded seals.
As Moore and Huntington (2008) have stated:
. . .the fitness of Arctic marine mammals will be influenced by change to the dynamic balance among sea ice’s effects on ecosystem structure and prey availability, as well as its role as a barrier or platform. In the ice-obligate category, the polar bear provides the clearest example: reductions in sea ice remove their hunting and resting platforms and likely reduce survivorship of ringed seals, their primary prey.
What is this ‘dynamic balance’ within which the Arctic is assumed to operate? It is a relatively modern notion (e.g. Wu and Loucks 1995) that eminent ecologist Daniel Botkin10 calls the ‘nature in balance’ concept:
The idea that nature can be restored to a single best condition is also part of a modern nature-myth, the belief in nature as a machine. According to this belief, developed in the nineteenth century, nature was like a watch or steam engine. It could be operated to run steadily. [See also Botkin’s short video summary of his book, “The Moon in the Nautilus Shell“]
Botkin also described a specific example of the fallacy of this out-of-date concept in this excerpt from his 1995 book (Botkin 1995:67–79.), in which he discusses modern grizzly bear populations compared to what explorers Lewis and Clark observed in the western United States in the early 1800s:
. . . this old view of nature also carried with it the belief that each creature existed at its carrying capacity – at the maximum number that could be sustained by its environment. Such a belief, while consistent with the idea of a perfect balance of nature, contradicts the inherent changeableness of the environment, which Lewis and Clark came to know all too well in their travels on the Missouri. And scientists know now that populations of grizzlies and other animals and plants are, like the Missouri River, always changing. There is no single ‘natural’ abundance.
There is a range of abundances, all of which are ‘natural’ in the sense that the population was at that level at some time during the past, prior to the effects of modern civilization.
The ecological concept of ‘stability’ is thus an idealised state, a theoretical condition that often does not apply to real animal populations and the environments they occupy in the real world (e.g. Huggett 2004:177). For example, amongst terrestrial mammals, drought or heavy snow can impact populations profoundly over the short term by decreasing critical food supplies or access to them (Gillson and Hoffman 2007; Hambler et al. 2005; Reimers 1982; Martinka 1967).
For polar bears and Arctic seals, sea ice has only been ‘stable’ in the sense that the habitat has been available, although variable in seasonal extent, over hundreds of thousands of years, through glacial ice ages and interglacial warm periods alike (Cronin et al. 2014; Polyak et al. 2010), but this is not how polar bear biologists use the term.
It is clear from the literature that polar bear specialists believe that polar bears, until recently, lived in ideal stable habitats that never varied and maintained stable or increasing population numbers that never declined due to natural environmental factors.
For example, Peacock et al. (2010), in a book chapter on polar bear conservation in Hudson Bay, provided a list of four reasons they thought sea ice habitat had not been explicitly protected over the last four decades and the fourth reason given was that:
…sea ice as a habitat has been considered stable in quantity and quality.
This assumption of sea ice as a naturally stable habitat over short time periods was also demonstrated in a 2014 BBC web essay, which quoted Dr. Steven Amstrup, a former polar bear biologist for the US Geological Survey and now ‘chief scientist’ for Polar Bears International, as saying the following:
It is important to remember that wild animals, if they have a stable healthy habitat, are a renewable resource and they can be managed sustainably. The problem with polar bears is that, right now, their habitat isn’t stable. [my bold]
Computer models used in 2008 to predict the likely response of polar bears to predicted sea ice changes over the next 100 years in support of listing polar bears as ‘threatened’ under the Endangered Species Act assumed population starting points were stable or increasing (Amstrup et al., 2007: 56; Amstrup et al. 2008, 2010; Amstrup 2011; Durner et al. 2009).
In addition, any negative habitat changes in any year were assumed to be due to unnatural increases in the length of the ice-free period in summer (i.e. due to human-caused global warming).
For example, in one of the nine USGS reports supporting the 2008 polar bear listing document, Hunter and colleagues (Hunter et al. 2007) divided five years worth of Southern Beaufort Sea population data into ‘good’ ice summers (2001–2003) and ‘bad’ ice summers (2004–2005) for the purpose of projecting future population responses to predicted summer sea ice declines (a ‘good’ ice year was defined as 50% ice coverage over the continental shelf, 125 days ice-free in preferred habitat).
They assumed that ‘bad’ ice summers were caused by unnatural (human-caused) influences while ‘good’ ice summers reflected normal, stable sea ice conditions, and thus should support a stable or increasing population. This assumption is reflected in a short statement from page 19 of their conclusions:
If conditions were to remain similar to 2001–2003, the population would
increase over the next 45–100 years.
Similarly, Stirling and colleagues (Stirling et al. 2007: 12), in their comparison of survival of cubs-of-the-year (bears born that spring; “COYs”) between Southern Beaufort and Northern Beaufort relative the amount of open water (similar to the ‘good’/‘bad’ summer ice conditions described above), had this to say;
. . .the marked reduction in survivorship for COYs noted in [the Southern Beaufort] in 2005 and 2006 (Regehr et al. 2006) was not observed in [the Northern Beaufort]. It is possible the drop in survivorship in [the south] was related to ecological changes resulting from several years of successively more extensive and extended open water (Regehr et al. 2007a) while, in comparison, ice conditions have remained relatively stable in [the north].
Regarding the prey of polar bears, the government document that laid out the argument for listing polar bears as threatened had this to say (US Fish &Wildlife Service 2008: 28239):
. . .polar bears are uniquely adapted to hunting on ice and need relatively large, stable seal populations to survive.
It is abundantly clear that the new ‘instablility’ apparently faced by polar bears and their prey refers to summer sea ice coverage alone, a habitat variability that can only be considered unprecedented if the wealth of data on spring sea ice habitat changes are ignored (see also Amstrup et al. 2007; Durner et al. 2009; Oakley et al. 2012; Wang et al. 2012).
All of these factors contribute to the conclusion that Arctic biologists stand out amongst colleagues doing research in other regions. For the most part, ecologists and biologists using ecological principles have moved beyond the simplistic, 1970s notion of r-selected versus K-selected population/habitat dynamics and most have developed more realistic methods of analysis (Gillson and Hoffman 2007; Reznick et al. 2002:1518; Wu and Loucks 1995).
In contrast, virtually all Arctic biologists remain idealistic in regards to their outmoded assumptions that sea ice is a naturally stable habitat over short time frames and that stable (or increasing) populations are the only natural trends expected for Arctic marine mammals. This attitude is especially disturbing because the strongest evidence refuting these assumptions comes from their own research.
Section 2. Evidence that sea ice is not a stable habitat
Although long-term records of sea ice thickness and snow depth over sea ice are not available on a global basis, observations from several regions have shown that sea ice across the Arctic is not a naturally stable habitat over decadal time scales, especially in winter through spring.
[Winter, January–March; Spring, April–June (Pilfold et al. 2015, in press)]
Due to their dependence on particular winter/spring sea ice conditions, Arctic seal populations have varied over time at a number of locations. As a consequence of the variability in seal numbers during the spring, when polar bears consume two-thirds of their annual prey, polar bear population numbers have likewise fluctuated over decades.
Survival and production of young for both seals and polar bears are inextricably linked to prevailing sea ice conditions from January through June (Stirling and Lunn 1997:176).
Eastern Beaufort
Take, for example, the Beaufort Sea – the area north of Alaska’s Arctic coast and west of the Central Canadian Archipelago. The USA and Canada share management of polar bears in this area. Too much thick spring ice in this region has had detrimental short term consequences for both ringed seal and polar bear numbers approximately every 10 years since at least the early 1960s. However, numbers have quickly recovered (Polar Bear News 2013; Stirling 2002:68).
The first of these population declines was described in the proceedings of the 1979 IUCN Polar Bear Specialist Group (PBSG) meeting (Stirling et al. 1980:52). The following statement from that report reveals the surprise that biologist Ian Stirling and colleagues felt at the instability they had witnessed in the field:
However, it was apparent from the studies of both polar bears and seals that their populations had undergone marked declines in numbers, productivity, and survival of young in 1974 and 1975. The decline apparently occurred because of natural causes that are not completely understood. Up until the present, the numbers of seals and bears in relation to the marine ecosystem have been regarded as being fairly static. This is the first time that major changes in numbers and reproductive parameters caused by natural influences have been documented in populations of arctic seals and polar bears. [my bold]
A few years later, Stirling and colleagues (Stirling et al. 1982:21) made the following statement about the same mid-1970s Southern Beaufort phenomenon, which impacted both ringed and bearded seals as well as polar bears, and shows they understood its implications for assessing environmental impacts:
Until recently, management of marine mammals in the Canadian Arctic, to the extent that they are managed at all, seems to have been based on the assumption that ecological conditions show little variability. Thus, once populations are counted or quotas are established, little change in population management takes place for long periods. The results of this study have clearly shown that ice conditions in the eastern Beaufort Sea can be highly variable, can influence other ecological parameters, and can cause changes in the distribution and abundance of ringed and bearded seals. We expect that similar variability will be documented in other areas of the Arctic when comparable studies have been completed.
What this means in terms of environmental assessment is that, because conditions are so variable, the consequences of possible man-made detrimental effects will vary depending on the status of the seal populations at the time. [my bold]
In 2008, Stirling and colleagues (Stirling et al. 2008:21; see also Smith 1987; Harwood et al. 2000, 2012) summarized the decadal nature of polar bear and ringed seal population fluctuations in relation to another episode of the phenomenon that occurred in 2004–2006. They noted that population declines of both seals and bears were always associated with thick spring ice conditions but recovered afterward:
The 1960s, 1970s, and 1980s each experienced a two- to three-year decline in seal productivity in the eastern Beaufort Sea and Amundsen Gulf, associated with heavy ice conditions, around mid-decade. Each was followed by a decline in polar bear reproduction and condition, after which both seal and bear populations recovered (Smith, 1987; Harwood et al., 2000; Stirling, 2002). The beginning of each of those three periods was associated with heavy ice conditions through the winter before the reproductive decline of the seals, followed by a late spring breakup. [my bold]
These authors also noted that the spring sea ice conditions in 2004–2006 (the most recent documented incident) were as bad as they had been in 1974, when Ian Stirling and Nick Lunn recorded some of the worst cases of starvation they had ever witnessed in the Arctic (Stirling and Lunn 1997:177):
. . . in the spring of 1974, when ringed seal pups first became scarce, we captured two very thin lone adult female polar bears that had nursed recently, from which we deduced they had already lost their litters. A third emaciated female was accompanied by two cubs which were so thin that one could barely walk. We have not seen females with cubs in this condition in the Beaufort Sea, or elsewhere in the Arctic, before or since. [my bold]
In the early 1970s, many ringed seals and polar bears were known to have moved out of the south-east Beaufort Sea and into the Chukchi Sea (Burns 1975:77), probably to escape thick spring ice conditions. This may have occurred in other thick-ice years but has not been documented.
The few ringed seal pups that were born in the south-eastern Beaufort during any thick-ice spring would have been hard for polar bears to locate within the deformed near-shore ice.
With few naïve, fat newborns to be had, predation-savvy adult seals and young subadults would have been the primary prey available to polar bears in heavy ice years, regardless of when they occurred, making successful seal hunts a much greater challenge than usual.
In their recently-published paper, Bromaghin and colleagues (Bromaghin et al. 2014:647) said this about the recurring sea ice phenomenon in the Southern Beaufort (SBS), one of which occurred during their study (2004–2006):
Extensive ice rubble and rafted floes during winter and spring are thought to have led to past declines in polar bear productivity in the SBS (Stirling et al. 1976, Amstrup et al. 1986, Stirling 2002), as well as during our investigation (Stirling et al. 2008).
Unfortunately, the precise amount of polar bear population decline during the 1974–1975 thick-ice events has never been calculated: the drop in numbers was only noted as being ‘marked’. The thick ice coverage in those years primarily affected ringed seals by reducing almost to zero the number of pups born (Harwood et al. 2000, 2012), resulting in the starvation and reduced survival of polar bears, especially cubs.
For the mid-2000s event, however, a newly formulated model (Bromaghin et al. 2013) was used to estimate that numbers of polar bears had declined by 25–50% (Bromaghin et al. 2014).
Since the response of ringed seals to conditions in 2004–2006 were virtually identical to 1974—1976, with virtually no seal pups born (Harwood et al. 2012), and lacking other information, it is reasonable to assume that in the mid-1970s, the polar bear population would have declined by similar amounts as in the mid-2000s, due to a combination of reduced cub survival and movement of bears out of the region.
In both periods, polar bear numbers rebounded to pre-thick ice levels within 3–6 years (Stirling and Lunn 1997: 176; Stirling 2002:68; and posts here, here, and here).
Other incidents in that region are not as well documented because polar bear research in the 1960s, 1980s, and 1990s was rather limited. However, ringed seal researchers noted that seal pup production fell to almost zero in 1963, 1984–1986, and in 1992—1994, suggesting that local polar bear populations probably faced starvation-related declines in survival or forced relocation pressure in those decades also (Stirling 2002; Stirling and Lunn 1997:174,176).
That makes a total of two well documented periods of significant declines in polar bear and ringed seal populations caused by natural changes in sea ice quality during the spring, with three additional events that may or may not have been as severe.
Stirling and Lunn (Stirling and Lunn 1997:176) concluded the following in their 1997 paper:
From the results presented above, it seems clear that the most critical factor affecting reproductive success, subsequent condition and probably survival of polar bears is the availability of ringed seal pups from about mid-April through to breakup sometime in July.
What they did not add is that availability of ringed seal pups in spring varied with spring sea ice conditions, especially ice thickness: that insight came a bit later (Stirling 2002; Stirling et al. 2008). Oddly, Stirling and others now link polar bear survival and population declines in the Southern Beaufort to sea ice conditions in late summer/fall (August to October)(Stirling and Derocher, 2012; Peacock et al. 2010; IUCN PBSG status table), although Nick Pilfold and colleagues (Pilfold et al. 2015, in press) recently conceded:
Because polar bears may acquire as much as two-thirds of their energetic intake in spring (Stirling and Øritsland 1995), a significant reduction in kill biomass could have population level consequences. Spring is the mating season for polar bears, and successfully mated females can avoid implantation or abort fetuses before denning if fat stores are inadequate (Derocher et al. 1992). Additionally, Bromaghin et al. (2015) reported lower survival rates between 2004–2006 for the southern Beaufort subpopulation and suggested measures of ice-availability were insufficient in explaining the decline. We suggest that poor hunting conditions in spring 2005–2006 could have negatively affected polar bear survival. [my bold]
Eastern Arctic: effects of the Atlantic Multidecadal Oscillation
Similar evidence of spring sea ice habitat instability comes from Hudson Bay and the Barents Sea. In Hudson Bay in the early and late 1990s, production and survival of ringed seal pups declined markedly (Chambellant et al. 2012; Stirling and Lunn 1997). The low number of pups in 1991 and 1992 was correlated with heavy spring sea-ice conditions and a much shortened open-water period. Ringed seal researcher Magaly Chambellant and colleagues blamed the thick spring ice on natural conditions and events (Chambellant et al. 2012:275):
The combined effect of a strong positive phase of the North Atlantic Oscillation [aka the Atlantic Multidecadal Oscillation, AMO (Wyatt and Curry 2014)] and a strong El Niño event in 1991–1992, and the eruption of Mount Pinatubo in June, 1991, created colder than average conditions in the eastern Arctic, including Hudson Bay, in the early 1990s.
Similarly to thick-spring-ice events in the Southern Beaufort, it was noted that some of the decline in seal populations in Hudson Bay during the 1990s may have been due to seals leaving the region during heavy ice years, although reduced pup survival in other years was correlated with spring snow depths being less than 32 cm.
Overall, Steve Ferguson and colleagues (Ferguson et al. 2005:127, Fig.2) summarised the variation in survival of ringed seals between the 1971 and 2001 this way:
The pooled survivorship curve for the western Hudson Bay sample (1999–2001) showed considerable annual variability. . .suggesting that survival of seals born in any given year was not constant. However, a decadal pattern emerged suggesting poor ringed seal recruitment, 1971–1978 (7 of 8 yr below average); better than average recruitment, 1979–1988 (8 of 10 yr); followed by poor recruitment in later years, 1989–2000 (8 of 12 yr). [my bold]
Due to the reliance of polar bears on abundant ringed seal pups in spring, survival of Western Hudson Bay cubs between spring and the following summer between 1980 and 1992 dropped to a low of 39% in 1991 (from 70–100% some years) when low seal pup production was noted due to thick spring ice.
Polar bear cub survival was also low (41–47%) in 1988—1990 (Derocher and Stirling 1996: 1247), when low snow depths reduced seal pup survival (Ferguson et al. 2005:127–128). In 1992, polar bears were reportedly in good condition when they left the ice in late summer. They had been feeding for about three weeks longer than average, apparently compensating for the lower availability of seal pups.
Negative polar bear cub survival clearly impacted overall population size: the largest sustained drops in western Hudson Bay occurred in the late 1980s and early 1990s (see Fig. 1) and again in 1999. These impacts were associated with documented declines in ringed seal pup production, which were blamed in turn on heavy ice conditions and/or low snow depths.
[Regarding the final population estimate for 2011, Lunn et al. (2013:18) noted that they calculated a new estimate for 2004 using the same method they used for 2011, which gave 742 (range 630–872) for 2004 and 806 (653-984) for 2011, indicating no decline since the last estimate was calculated in 2004 by Regehr et al. (2007a)]
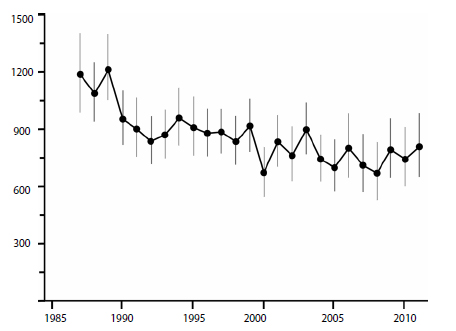
Figure 1: Polar bear population in W. Hudson Bay. Redrawn from Lunn et al. 2013:40. Click to enlarge.
The 30-year trend towards earlier breakup dates in western Hudson Bay does correlate with overall population declines since the 1980s but rarely do specific early breakup dates correlate with short-term declines in population size. For example, breakup was apparently early in 1990 (Fig. 2) but, as noted above, low ringed seal production that year was blamed on shallow winter snow depths and there was an associated decline in polar bear numbers (Fig. 1).
In 2003, breakup was very early (Fig. 2) and was associated with only a modest decline in polar bear numbers over the next few years (Fig.1). Lunn and colleagues (Lunn et al. 2013:15, 21) found no trend in breakup or freeze-up dates between 2001–2010 – breakup dates were not getting earlier over time nor were freeze-up dates getting later.
[Similarly, according to Stirling et al. 2004 (Fig. 3, ‘Area 5’), breakup in WHB near Churchill was also quite early in 1976 (ca. June 16) and again in 1977 (ca. June 9) but number of bears counted did not show an associated decline (Fig. 4b): the lowest number of bears counted after 1975 occurred in 1979]
In contrast, seal pup production and polar bear cub survival were both low in 1991 (Fig. 2), but both were blamed on the relatively late breakup of sea ice that year and its associated short open-water season.
Unfortunately, spring snow depths over sea ice and spring ice thickness for Hudson Bay do not appear to be reported independently of ringed seal research reports, which means polar bear biologists are lacking a long-term, detailed record of these critical winter and spring sea ice habitat characteristics to include in their analyses and predictive models.
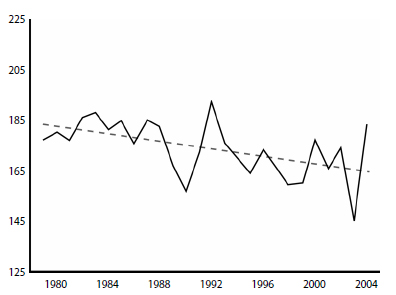
Figure 2: Breakup dates for W. Hudson Bay. The figure uses the 50% ice cover threshold for ‘breakup’ and Julian calendar days. Dashed line is the authors’ trend line fit by linear regression. Redrawn from Stirling and Parkinson (2006: 265). Click to enlarge.
Multidecadal variability in winter sea ice thickness has also been reported from the late 1700s to the mid-1900s by Danish biologist Christian Vibe (Vibe 1965:17–24) for northwest Greenland (i.e. Baffin Bay) as well as the north coast of East Greenland (Fig. 3). The variability correlated strongly with changes in availability of ringed seals and polar bears.
Bears and seals apparently moved out of areas when thick winter/spring ice conditions dominated but returned when normal ice conditions resumed.
In contrast, around the Svalbard Archipelago in the eastern Arctic, the condition of polar bears has been linked to the relative coverage of sea ice in spring, which varies naturally with the state of the AMO. In this case, the variation has not been in ice thickness, but ice extent (Fig. 3).
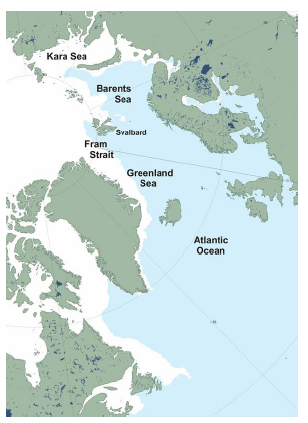
Figure 3: Sea ice extent around Greenland and the Barents Sea at 8 March 2014. NSIDC MASIE sea ice product. Click to enlarge.
At the end of February 2014, the US National Snow and Ice Data Center noted that ice in the Barents Sea was below the seasonal average due to natural variations driven by the AMO, a factor that had been mentioned previously in relation to Hudson Bay ice variation. The AMO (Fig. 4), according to one source (Miles et al. 2014:462):
. . . is a coherent pattern of basin-wide sea surface temperature (SST) variations with a period of roughly 60–90 years. . . Paleoenvironmental studies suggest that the AMO has persisted through previous centuries. . .and even millennia. . .
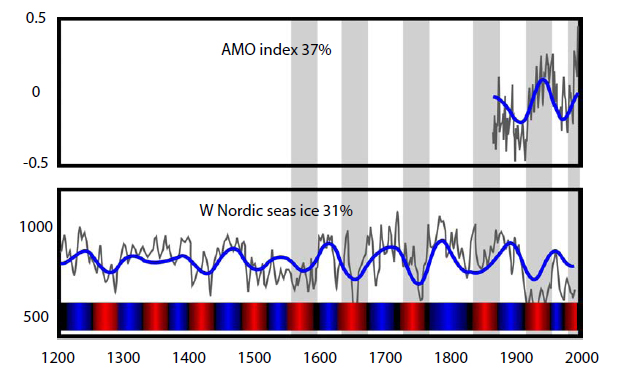
Figure 4: Persistent multidecadal fluctuations in sea ice linked to the AMO. Top, AMO modern index for the North Atlantic sea surface temperature anomaly; bottom, the extended record going back 400 years for sea ice extent (based on proxy reconstructions) for the East Greenland Sea. The color bar indicates periods with reduced ice (red) and periods with increased ice (blue) inferred from the wavelet-filtered signal. The reduced ice periods are seen to correspond to warm AMO periods. Source: Redrawn from Miles et al. (2014). Click to enlarge.
Norwegian biologists Jon Aars and Magnus Andersen of the Norwegian Polar Institute have pointed out in their online polar bear research updates that around Svalbard the body condition of adult males, number of females with cubs-of-the-year, and litter size varied with the AMO and sea-ice levels in spring (April–June).
Miles and colleagues concluded, regarding the AMO’s influence on sea ice (Miles et al. 2014:468):
The pervasive multidecadal variability in observed sea ice is here not
considered to represent truly oscillatory cycles but rather irregular, broadly multidecadal fluctuations betweenwarmer (colder) periods with less (more) ice that are related to [Atlantic multidecadal variability].
Polar bear biologist Andrew Derocher looked for correlations of climate with polar bear body condition in spring (1988–2002) in a study of Svalbard bears. He focused on the Arctic Oscillation (AO)(Ambaum et al. 2001) rather than the AMO, but in comparing his Barents Sea results to those from similar studies in western Hudson Bay (Derocher 2005:273), he stated that:
The stronger correlation [found] between the Arctic Oscillation index in the spring for both females and males suggests that climate influences affect polar bears more during this period. [my bold]
Canadian Arctic Archipelago
Another example of the effects of variable spring snow depth and sea ice on both ringed seal and polar bear survival comes from the work of Mike Hammill and Tom Smith (Hammill and Smith 1991:125), who studied polar bear predation on ringed seals in the Lancaster Sound region of the central Canadian Arctic (known as Barrow Strait).
They encountered significant variability in sea ice conditions and snow cover over a three-year period in the mid-1980s. Working in March and April (late winter/early spring), they noted:
. . . in 1986, extensive open water conditions forced us to survey different plots. . .
They also recorded that mean snow depth declined from 23 cm in 1984 to 10 cm in 1985. The decline in snow depth between years coincided with an increase in predation success by polar bears on newborn ringed seals, suggesting that deep spring snow offered ringed seals more protection from polar bear attack.
As to the importance of spring as a feeding period for polar bears, they had this to say (Hammill and Smith 1991:132):
Prior to late April, we found little evidence of predation in Barrow Strait. In late spring, polar bears enter a period of intense feeding (Stirling and McEwan 1975, Ramsay and Stirling 1988), which begins with the onset of the ringed seal pupping season.. .Feeding on young seals continues throughout the spring and early summer as bears replenish depleted fat reserves. After ice breakup, bears move ashore and begin another period of little feeding (Stirling and McEwan 1975, Ramsay and Stirling 1988). [my bold]
Ian Stirling and Andrew Derocher reiterated this essential point in their recent summary paper (Stirling and Derocher 2012:2698):
Stirling & Øritsland (1995) estimated that in most areas, polar bears likely accumulate two thirds or more of the energy they will need for the entire year during the late spring and early summer before breakup. The long open water season in much of the Arctic necessitates accumulation of fat reserves. [my bold] [See also Stirling 1974]
In other words, the spring feeding period is critical for polar bears because they eat so little during the rest of the year.
A review of the literature shows little documented evidence for polar bears hunting seals successfully in late summer (Derocher et al. 2002; Hammill and Smith 1991; Stirling 1974; Stirling and Øritsland 1995): in most regions they do not eat much, regardless of whether summers are spent on shore or on the sea ice. In fact, many of the effects of a longer open-water period have been unexpectedly positive for polar bears.
For example, polar bear condition and reproduction in the Chukchi Sea (between Russia and Alaska) was much better over a recent period with a long open-water season than it was in the 1980s, when there was a short open water season. Ringed seals flourished during the recent long open-water season: they had a longer time to feed and consequently reproduced well.
Similarly, in the southern Davis Strait (off Labrador), polar bears have flourished despite longer ice-free seasons in recent years because harp seal numbers increased exponentially over the same period (due to collapse of commercial seal hunting).
In both cases, more healthy seals meant more food for polar bears in spring.
Polar bears that have plenty of seals to eat in spring are fat by early summer, and that fat serves as an essential energy buffer over the summer months, when bears in most regions fast or eat very little.
While the timing of ice breakup or extent of summer sea ice is also variable, well-fed bears are prepared for a summer fast of variable length.
In conclusion, it is critical to note that in the Barents Sea/East Greenland region we have evidence of marked natural variability in winter/spring sea ice conditions going back at least 400 years.
Variability in spring sea ice and snow conditions has been shown to affect polar bear and ringed seal condition and reproduction over the short term, and may initiate movements of animals between regions.
The most deadly of these variations for both species appears to be thick spring ice, although shallow snow cover over spring ice may be nearly as detrimental. As a result, both polar bear and ringed seal population size and reproduction have varied considerably in several regions over the last 20–60 years.
So far, few of the marked declines documented in polar bear populations
over the last 40 years discussed here can be plausibly blamed on reduced summer sea ice – apparent correlations are spurious or coincidental.
Therefore, the evidence is strong that the ‘natural’ state for Arctic sea ice habitat in the winter and spring is not stable over short time frames. Consequently, neither are polar bear and ringed seal population numbers.
Section 3. Sea ice and Arctic marine mammal
conservation
Polar bear conservation started several decades before other Arctic marine mammals, with the exception of whales (the International Whaling Commission was established in 1946).
In the late 1950s, Russia banned all polar bear hunting. By the late 1960s, it was recognized internationally that polar bear populations around the Arctic had been seriously depleted by wanton slaughter and unregulated hunting and that they were at risk of extinction unless coordinated steps were taken to halt the process.
In 1973, an international treaty was signed by Arctic nations [Canada, Russia, Denmark for Greenland, Norway, and the United States of America] to ensure the protection of polar bears against the over-hunting that threatened their survival.
The PBSG, formed in conjunction with that treaty, had from its inception a unique international authority to inform Arctic policymakers regarding the status of polar bear research and the relative health of polar bear populations. It took until 1988 for the IUCN to gather enough information to formally classify polar bears as ‘vulnerable’ (equivalent to the term ‘threatened’ under other schemes).
Just eight years later in 1996, polar bear population numbers had sufficiently recovered for the status to be boosted to ‘least concern.’ International protection from over-hunting had done its job: the polar bear had been saved.
Fast-forward to 2005. That year, the PBSG recommended that the IUCN return polar bears to ‘vulnerable’ status on the basis of future risks due to predictions of summer sea ice declines associated with man-made global warming.
The justifications offered for this recommendation were weak, to say the least. Little to no mathematically supported data or model results had been provided. For example, a glossy overview document (with no references to original research and sea ice projection models based on only 19 years of data – less than a single period for what is considered to be ‘climate’), was all that was offered regarding future polar bear habitat changes (Hassol 2004).
The professional opinion of polar bear specialists, namely that a population reduction of more than 30% within the next 35–50 years was foreseen (Aars et al. 2006:31), was apparently enough for the IUCN to accept the PBSG recommendation. As a consequence, in 2006, the polar bear was upgraded to ‘vulnerable’ on the IUCN Red List.
Using a similar approach, in 2008 the US Fish and Wildlife Service listed polar bears as ‘threatened’ under the Endangered Species Act, a status equivalent to the IUCN category of ‘vulnerable’ (US Fish and Wildlife Service 2008). This was the first time that a predicted threat based on computer models had been accepted under the terms of the Act as evidence for a ‘threatened’ listing.
Even though the USA possessed the smallest proportion of the world’s polar bear habitat, the Act pronounced a global endangerment finding. Supported by eight internal government reports commissioned by the US Fish and Wildlife Service and prepared by the US Geological Survey rather than peer-reviewed academic papers, the justification for this listing hinged on what are called ‘Bayesian Network’ model results.
The models relied upon the opinions of a single expert regarding how polar bears might respond to predicted sea ice changes. The same complex Bayesian Network model approach was recently used to assess walrus responses to predicted sea ice declines (Jay et al. 2011:1068; Oakley et al. 2012:4), using four expert opinions.
However, the use of such complex but data-poor Bayesian Network models has been strongly criticized for this purpose (Akçakaya et al. 2006; Karl et al. 2012) a point that will be discussed in more detail below.
The 2007 polar bear model has since been revised to include the expert opinions of four additional biologists (Atwood et al. 2014), but still does not include any population estimates, any discussion of naturally-occurring variations in population size, or changes in polar bear health due to naturally occurring shifts in sea-ice conditions, particularly in the critical spring season.
The complex ‘second generation’ model produced in 2014 by Atwood and colleagues assumes the only sea ice change that ‘threatens’ polar bear health or population size is the predicted decline in summer ice extent blamed on human-caused global warming. The well-documented variations in spring ice thickness in the Southern Beaufort, and their associated repercussions on seal and polar bear populations, have been glossed over in favor of spurious correlations with summer-ice declines (Fig. 5).

Figure 5: Ice-free days in the southern Beaufort Sea. Number of ice-free days. Solid circles are the years used in the polar bear population survival vs. sea ice study. Note that ‘good’ summer ice years are those below 125 ice-free days and ‘bad’ summer ice years (those blamed on global warming) are above 125 day. Redrawn from Regehr et al. 2007a:26. Click to enlarge.
The focus of American biologists on opinion-heavy but data-light Bayesian Network models, based on the fallacy that sea ice is a stable habitat (see Section 2), has led to a peculiar international conservation bias: the USA, alone amongst all Arctic nations, has listed bearded seals, ringed seals, and spotted seals as ‘threatened’ based on predicted sea ice declines due to global warming (US Fish and Wildlife Service 2012a and 2012b).
A similar listing for the Pacific walrus is pending (US Fish and Wildlife Service 2011). However, a court-ordered review of the bearded seal ESA listing is underway and a petition to list the Pacific ribbon seal under the same ‘future threats’ criteria was rejected.
The USA has been uniquely aggressive amongst Arctic nations in assigning ‘threatened’ status to Arctic seals based on predicted population declines blamed on human-caused global warming. Most other countries, as well as the IUCN, list Arctic seals as either being of ‘least concern’ or have not assessed them at all.
There are indications that this situation may change in the next few years, at least for the IUCN Red List status of polar bears. The first-generation predictive model used to support the US listing of polar bears as ‘threatened’ (Amstrup et al. 2008) recently came under heavy criticism from the chairman of the IUCN Standards and Petitions subcommittee, a biological modeling expert.
Almost certainly because of those concerns, the next IUCN polar bear assessment, due in June 2015, will have to meet strict new IUCN standards for using predicted climate change threats as criteria for recommending a ‘vulnerable’ or ‘endangered’ status (IUCN Standards and Petitions Subcommittee 2014). However, none of those new IUCN standards acknowledge that sea ice is a naturally variable habitat over short timescales.
That suggests the next IUCN Red List assessment for polar bears will be just as biologically flawed as the American judgment.
A close examination of the models developed to predict future population changes for polar bears, walrus, and ice-dependent seals reveals they all assume the fallacy that under ‘natural’ conditions – without human-caused global warming – sea ice provides a stable habitat.
The models also assume, of course, that populations living in such stable habitats should grow or remain stable. As a consequence, the computer models that project population risks over the next 30–45 years focus on the end of summer (Overland and Wang 2013; Swart et al. 2015), the only season for which recent declines in sea ice have been pronounced (see Fig. 6).
Projected declines in winter ice are insignificant, and those for spring ice are generally slight (see Durner et al. 2009: 44 for a general Arctic projection and Wang et al. 2012 regarding Chukchi Sea projections).
For example, Amstrup et al. (2007:9) stated:
. . . all GCMs project extensive winter sea ice through the end of the 21st century in most ecoregions (Durner et al. 2009).[Note that Steven Amstrup was the USGS biologist whose ‘expert opinion’ was used in the Bayesian Network models; he is employed as ‘chief scientist’ at the conservation organization Polar Bears International]
Negative effects on populations of short-term natural variations in spring sea ice or spring snow cover on ice have been entirely ignored in these predictions and the focus, both in academic reports and media stories, has been on summer ice extent.
More disturbing is that population declines that were clearly caused by thick sea ice conditions in spring are now blamed on summer sea-ice declines. I repeat, since this is such a critical point, population declines that were clearly caused by thick sea ice conditions in spring are now blamed on summer sea-ice declines. These claims are based on spurious statistical correlations, and are used as ‘evidence’ that the predicted declines of sea ice in the future will have more pronounced effects.
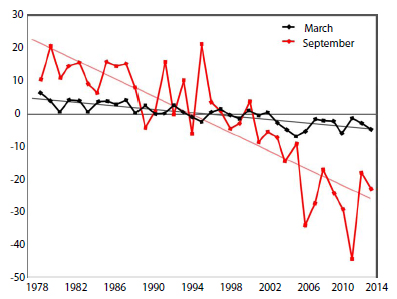
Figure 6: September and March sea-ice declines. Source: NOAA, Arctic Report Card 2014. Click to enlarge.
Section 4. Consequences of recent sea ice declines in summer
Surprisingly, negative changes to polar bear and ringed seal populations in response to summer sea-ice declines since 2006 have so far not materialized, despite many confident pronouncements based on expert opinion (e.g., Stirling and Derocher 2012; Obbard et al. 2010; Laidre et al. 2008).
Unfortunately, the Bayesian models that were published in 2007 to support the ESA ‘threatened’ listing generated only ‘probabilities of extinction’ at 2045, 2075, and 2100 (Amstrup et al. 2007:92; Durner et al. 2009). These dates were too far in the future, and the concept too mathematically vague, to allow the models to be tested against observations.
The few marked population declines that have been documented since the 1980s, such as in the Southern Beaufort and Western Hudson Bay, were actually associated with short-term episodes of thick spring ice and/or low spring snow depth rather than long-term declines in summer ice coverage.
Only one incident of summer sea ice decline – the early breakup of sea ice in Western Hudson Bay in 2003 – was clearly associated with a documented drop in polar bear numbers, although the decline was small compared to earlier ones associated with variable spring habitat conditions.
Contrary to predictions, reduced summer ice in the Chukchi Sea in the 21st century has proven to be a huge benefit to ringed seals (Crawford and Quakenbush 2013; Rode et al. 2014; Rode et al. 2013) because they do most of their feeding during the ice-free season: more food for ringed seals in summer has meant more pups the following spring for hungry polar bears.
Similarly, marked increases in the ice-free period in the southern Davis Strait have been offset by abundant harp seal in the spring – so much so that the population has increased despite dramatic summer sea ice declines (Peacock et al. 2013; Rode et al. 2012).
Most recently, reports on fieldwork into 2013 suggests that Southern Beaufort polar bear populations have not been negatively affected by the remarkable decline of sea ice in late summer of 2012 (Fig. 6). The western Arctic was particularly severely affected in 2012, and saw the highest number of ice-free days since 1987, as shown in Fig. 5 (Rode et al. 2014).
Reduced summer ice has apparently not been accompanied by reduced spring snow depth over sea ice in any region, as biologists expected (Stirling and Derocher 2012; Crawford and Quakenbush 2013; Rode et al. 2013) – no such snow depth declines have been reported.
While it is true that there have been moderate increases in the length of the ice-free season in regions of the Arctic where the ice usually melts completely in the summer – for example Hudson Bay and the Davis Strait – this has so far changed gradually and polar bear populations are either stable or increasing (Cherry et al. 2013; Obbard et al. 2006, 2007, 2010, 2013; Lunn et al. 2013).
The most recent research calculated breakup dates in relation to ice-cover levels corresponding to when polar bears actually left the ice in summer and returned to it in the fall, rather than an arbitrary 50% cut-off (Cherry et al. 2013). It found that breakup dates for Western Hudson Bay over the last 20 years have so far had little or no impact on the critical feeding period. In other words, breakup has so far not come before mid-June, which means most bears have come onshore in early July or later, although some individuals may come ashore before then.
And as noted previously, it is apparent that no matter where polar bears spend the summer – on shore or on the ice – they generally consume little during the open-water season. Bears that have hunted successfully between March and June under suitable spring conditions will have stored enough fat to see them through a summer fast of 2–6 months, whether they spend that time on land or out on the sea ice.
Section 5. Conclusions
The notion that declines in summer sea ice are the most serious threat to Arctic marine mammals is based on the biological fallacy that under natural conditions Arctic sea ice is a stable habitat and leads to stable or increasing populations, and that summer sea ice stability is necessary to prevent extinctions.
This flawed ecological concept has led to the present pessimistic attitude of most polar bear biologists and to the formal, legal listing by the USA of polar bears, walrus, ringed seals, bearded seals and spotted seals as ‘threatened’. The sea ice constancy fallacy was taught to leading polar bear experts years ago and accepted as fact, but to their credit – even though they believed sea ice should be a stable habitat, even over short timescales – they spent years collecting evidence to the contrary.
Eventually, the concept of human caused global warming provided a rationale for deferring to what they had been taught over what they had personally discovered, and they convinced themselves that reduced summer ice coverage could potentially pose as large a threat to polar bear and Arctic seal survival as reckless overhunting.
However, there is well documented evidence that natural changes in sea ice during the critical spring period over the last 60 years have been profound and resulted in marked declines followed by marked increases in polar bear and Arctic seal numbers.
Rather than climate change leading to extinction or near-extinction of these species, they appear to persist well, despite such environmental variation, even during the critical spring period. Historical documents and paleoclimatic research suggests similar changes have occurred for at least the last few hundred years.
Since the 1960s, thick spring ice has been more prevalent, and affected more Arctic regions, than scarce summer ice. Most important, since the thick ice phenomenon is caused by natural forces rather than presumed man-made ones – and will almost certainly continue into the future – conservation policies currently in place cannot prevent populations from fluctuating.
Like other temperate marine mammal species, such as the humpback whale, polar bears are thriving in a world that has finally protected them from relentless slaughter. Their populations are now free to respond to natural variations in sea-ice habitat and food supply, as they have done for hundreds of thousands of years.
Pretending that natural fluctuations in populations don’t happen in the Arctic won’t prevent them from happening.
The fallacy of sea ice stability is biologically unsupportable, and means that predictive population models based on this premise are not using the best available science. Some would argue they are not based on accepted scientific practice at all.
The IUCN and the US government, in accepting predicted sea-ice changes as valid threats to species survival, have allowed a myth to masquerade as 21st century science. A model built on a flawed premise is itself flawed.
Until such time as predictive models are modified to reflect the reality of Arctic sea ice as a naturally variable habitat – and populations of polar bears and their prey acknowledged as highly variable over time due to natural habitat changes in spring – the notion that the population impact of one particular kind of habitat change (‘human-caused’ summer sea-ice coverage) can be predicted within a biologically meaningful time frame must be abandoned.
Given what we now know about the animals and their naturally changing habitat, it is time to concede that data do not support predictions that polar bears, walrus, and Arctic seals are threatened with extinction due to habitat instability.
Bibliography
Aars, J., Lunn, N.J. and Derocher, A.E. (eds.) 2006. Polar Bears: Proceedings of the 14th Working Meeting of the IUCN/SSC Polar Bear Specialist Group, 20–24 June 2005, Seattle,Washington, USA. Occasional Paper of the IUCN Species Survival Commission 32. IUCN, Gland (Switzerland) and Cambridge (UK).
Adler, J.H. 2008. An animal to save the world: climate change and the polar bear. The New Atlantis 21 (summer): 111–115.
Akçakaya, H.R., Butchart, S.H.M.,Mace, G.M., Stuart, S.N. andHilton-Taylor, C. 2006. Global Change Biology 12: 2037–2043.
Ambaum, M.H.P, Hoskins, B.J. and Stephenson, D.B. 2001. Arctic Oscillation or North Atlantic Oscillation? Journal of Climate 14: 3495–3507.
Amstrup, S.C. 2011. Polar bears and climate change: certainties, uncertainties, and hope in awarmingworld. In: R.T.Watson, T.J. Cade, M. Fuller, G. Hunt and E. Potapov (eds.), Gyrfalcons and Ptarmigan in a Changing World, Volume 1. The Peregrine Fund, Boise, Idaho.
Amstrup, S.C., Stirling, I. and Lentfer, J.W. 1986. Past and present status of polar bears in Alaska. Wildlife Society Bulletin 14: 241–254.
Amstrup, S.C.,Marcot, B.G. andDouglas,D.C. 2007. Forecasting the rangewide status of polar bears at selected times in the 21st century. Administrative Report, US Geological Survey. Reston, Virginia.
Amstrup, S.C.,Marcot, B.G.,Douglas,D.C. 2008. A Bayesian network modeling approach to forecasting the 21st century worldwide status of polar bears. Pp. 213–268 in Arctic Sea Ice Decline: Observations, Projections, Mechanisms, and Implications, E.T. DeWeaver, C.M. Bitz and L.B. Tremblay (eds.). Geophysical Monograph 180. American Geophysical Union, Washington, D.C.
Amstrup, S.C., DeWeaver, E.T., Douglas, D.C., Marcot, B.G., Durner, G.M., Bitz, C.M. and Bailey, D.A. 2010. Greenhouse gas mitigation can reduce sea-ice loss and increase polar bear persistence. Nature 468: 955–958.
Anthony, R.G. 1976. Influence of drought on diets and numbers of desert deer. Journal of Wildlife Management 40(1):140–144.
Atwood, T.C.,Marcot, B.G.,Douglas,D.C., Amstrup, S.C., Rode, K.D.,Durner, G.M. and Bromaghin, J.F. 2014. Evaluating and ranking threats to the long-termpersistence of polar bears. USGS Open-File Report 2014–1254.
Botkin, D.B. 1995. Our Natural History: The Lessons of Lewis and Clark, Putnam, NY.
Burns, J.J. 1975. Quarterly report. In: Environmental Assessment of the Alaskan Continental Shelf, Principal Investigators’ Reports, July-September 1975. Boulder.
Bromaghin, J.F., McDonald, T.L., Stirling, I., Derocher, A.E., Richardson, E.S., Rehehr, E.V., Douglas, D.C., Durner, G.M., Atwood, T. and Amstrup, S.C. 2015. Polar bear population dynamics in the southern Beaufort Sea during a period of sea ice decline. Ecological Applications 25(3):634–651.
Chambellant, M., Stirling, I., Gough, W.A. and Ferguson, S.H. 2012. Temporal variations in Hudson Bay ringed seal (Phoca hispida ) life-history parameters in relation to environment. Journal of Mammalogy 93: 267–281.
Cherry, S.G., Derocher, A.E., Thiemann, G.W., Lunn, N.J. 2013. Migration phenology and seasonal fidelity of an Arctic marine predator in relation to sea ice dynamics. Journal of Animal Ecology 82: 912–921.
Crawford, J. and Quakenbush, L. 2013. Ringed seals and climate change: early predictions versus recent observations in Alaska. Oral presentation by Justin Crawfort, 28th LowellWakefield Fisheries Symposium, March 26–29, Anchorage, AK. http://seagrant.uaf.edu/conferences/2013/wakefield-arctic-ecosystems/program.php.
Cronin, M.A., Rincon, G.,Meredith, R.W.,MacNeil, M.D., Islas-Trejo, A., Cañovas, A. and Medrano, J. F. 2014. Molecular phylogeny and SNP variation of polar bears (Ursus maritimus ), brownbears (U. arctos ), and black bears (U. americanus ) derived from genome sequences. Journal of Heredity 105: 312–323.
Derocher, A.E. 2005. Population ecology of polar bears at Svalbard, Norway. Population Ecology 47: 267–275.
Derocher, A.E. and Stirling, I. 1996. Aspects of survival in juvenile polar bears. Canadian Journal of Zoology 73: 1246–1252.
Derocher, A.E., Stirling, I. and Andriashek, D. 1992. Pregnancy rates and progesterone levels of polar bears in western Hudson Bay. Canadian Journal of Zoology 70: 561–566.
Derocher, A.E.,Wiig,Ø. andAndersen, M. 2002. Diet composition of polar bears in Svalbard and thewestern Barents Sea. Polar Biology 25 (6): 448–452.
Derocher, A.E., Lunn, N.J. and Stirling, I. 2004. Polar bears in a warming climate. Integrative and Comparative Biology 44: 163–176.
Durner, G.M., Douglas, D.C., Nielson, R.M., Amstrup, S.C., McDonald, T.L. and 12 others. 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79: 25–58.
Ferguson, S.H., Stirling, I. and McLoughlin, P. 2005. Climate change and ringed seal (Phoca hispida ) recruitment in Western Hudson Bay. Marine Mammal Science 21: 121–135.
Gillson, L. and Hoffman, M.T. 2007. Rangeland ecology in a changing world. Science 315: 53–54.
Gray, S., Graumlich, L., Betancourt, J. and Pederson, G. 2004. A tree-ring based reconstruction of the AtlanticMultidecadal Oscillation since 1567 A.D. Geophysical Research Letters 31: L12205, doi:10.1029/2004GL019932.
Hambler, C., Henderson, P.A. and Speight, M.R. 2005. Elephants, ecology, and nonequilibrium? Science 307: 673.
Hammill, M.O. and Smith T.G. 1991. The role of predation in the ecology of the ringed seal in BarrowStrait, Northwest Territories, Canada. Marine Mammal Science 7: 123–135.
Harwood, L.A., Smith, T.G. and Melling, H. 2000. Variation in reproduction and body condition of the ringed seal (Phoca hispida ) in western Prince Albert Sound, NT, Canada, as assessed through a harvest-based sampling program. Arctic 53(4): 422 – 431.
Harwood, L.A., Smith, T.G., Melling, H., Alikamik, J. and Kingsley, M.C.S. 2012. Ringed seals and sea ice in Canada’swestern Arctic: harvest-based monitoring 1992–2011. Arctic 65: 377–390.
Hassol, S.J. 2004. Impacts of a Warming Arctic : Arctic Climate Impact Assessment. Cambridge University Press, Cambridge UK.
Huggett, R.J. 2004. Fundamentals of Biogeography, 2nd edn. Routledge, Abingdon.
Hunter, C.M., Caswell, H., Runge, M.C., Regehr, E.V., Amstrup, S.C. and Stirling, I. 2007. Polar bears in the southern Beaufort Sea II: Demography and population growth in relation to sea ice conditions. Administrative Report, US Geological Survey. Reston, Virginia.
IUCN Standards and Petitions Subcommittee. 2014. Guidelines for using the IUCN Red List Categories and Criteria. Version 11. Prepared by the Standards and Petitions Subcommittee.
Jay, C.V., Marcot, B.G. and Douglas, D.C. 2011. Projected status of the Pacific walrus (Odobenus rosmarus divergens ) in the twenty-first century. Polar Biology 34: 1065–1084.
Karl, S.A., Toonen, R.J., Grant, W.S. and Bowen, B.W. 2012. Common misconceptions in molecular ecology: echoes of the modern synthesis. Molecular Ecology 21: 4171–4189.
Knudsen, M.F., Seidenkrantz, M.-S., Jacobsen, B.H. and Kuijpers, A. 2011. Tracking theAtlanticMultidecadal Oscillation through the last 8,000 years. Nature Communications 2: 178, doi:10.1038/ncomms1186.
Laidre, K. L., Stirling, I., Lowry, L. F., Wiig, Ø., Heide-Jørgensen, M. P. and Ferguson, S.H. 2008. Quantifying the sensitivity of arctic marine mammals to climate-induced habitat change. Ecological Applications 18(2, Suppl.): S97–S125.
Lunn, N.J., Regehr, E.V., Servanty, S., Converse, S., Richardson, E. and Stirling, I. 2013. Demography and population assessment of polar bears in Western Hudson Bay, Canada. Environment Canada Research Report. 26 November 2013.
Martinka, C.J. 1967. Mortality of northern Montana pronghorns in a severe winter. Journal ofWildlife Management 31(1): 159–164.
Miles, M.W., Divine, D.V., Furevik, T., Jansen, E., Moros, M. and Ogilvie, A.E.J. 2014. A signal of persistent Atlantic multidecadal variability in Arctic sea ice.Geophysical Research Letters 41, doi:10.1002/2013GL058084.
Moore, S.E. and Huntington, H.P. 2008. Arctic marine mammals and climate change: impacts and resilience. EcologicalApplications 18(2, Suppl): S157–S165.
Oakley, K., Whalen, M., Douglas, D., Udevitz, M., Atwood, T. and Jay, C. 2012. Changing Arctic Ecosystems: Polar bear and walrus response to the rapid decline in Arctic sea ice. U.S. Geological Survey Fact Sheet 2012–3131.
Obbard, M.E., Cattet, M.R.L., Moody, T., Walton, L.R., Potter, D., Inglis, J. and Chenier, C. 2006. Temporal trends in the body condition of southern Hudson Bay polar bears. Ontario Ministry of Natural Resources, Applied Research and Development Branch, Sault Ste, Marie, Canada. Climate Change Research Information Note 3.
Obbard, M.E., McDonald, T.L., Howe, E.J., Regehr, E.V. and Richardson, E.S. 2007. Polar bear population status in southern Hudson Bay, Canada. Administrative Report, U.S. Department of the Interior- U.S. Geological Survey, Reston, VA.
Obbard, M.E., Theimann, G.W., Peacock, E. and DeBryn, T.D. (eds) 2010. Polar Bears: Proceedings of the 15th meeting of the Polar Bear Specialists Group IUCN/SSC, 29June-3 July, 2009, Copenhagen,Denmark. Gland, Switzerland and Cambridge UK, IUCN. http://pbsg.npolar.no/en/meetings/
Obbard, M.E., Middel, K.R., Stapleton, S., Thibault, I., Brodeur, V. and Jutras, C. 2013. Estimating abundance of the Southern Hudson Bay polar bears subpopulation using aerial surveys, 2011 and 2012. Ontario Ministry of Natural Resources,Wildlife Research andMonitoring Section, Science and Research Branch, Wildlife Research Series 2013–01. Peterborough, Ontario.
Overland, J.E. andWang, M. 2013. When will the summer Arctic be nearly sea ice-free? Geophysical Research Letters 40: 2097–2101.
Peacock, E., Derocher, A.E., Lunn, N.J. and Obbard, M.E. 2010. Polar bear ecology and management in Hudson Bay in the face of climate change. In: A Little Less Arctic: Top Predators in the World’s Largest Northern Inland Sea, Hudson Bay. S.H. Ferguson, L.L. Loseto and M.L. Mallory (eds). Springer.
Peacock, E., Taylor, M.K., Laake, J. and Stirling, I. 2013. Population ecology of polar bears in Davis Strait, Canada and Greenland. Journal of Wildlife Management 77: 463–476.
Pilfold, N. W., Derocher, A. E., Stirling, I. and Richardson, E. 2015 in press. Multi-temporal factors influence predation for polar bears in a changing climate. Oikos. doi: 10.1111/oik.02000
Polar Bear News. 2013. Polar bear newsletter of the US Fish & Wildlife Service, Anchorage, Alaska.
Polyak, L., Alley, R.B., Andrews, J.T. et al. 2010. History of sea ice in the Arctic. Quaternary Science Reviews 29: 1757–1778.
Pianka, E.R. 1970. On r and K selection. American Naturalist 104 (940): 592–597.
Ramsay, M.A. and Stirling, I. 1988. Reproductive biology and ecology of female polar bears (Ursus maritimus ). Journal of Zoology, London 214: 601–634.
Regehr, E.V. and Amstrup, S.C. 2006. Polar bear population status in the Southern Beaufort Sea. Open-File Report 2006–1337, US Geological Survey. Reston, Virginia.
Regehr, E.V., Hunter, C.M., Caswell, H., Amstrup, S.C. and Stirling, I. 2007a. Polar bears in the southern Beaufort Sea I: survival and breeding in relation to sea ice conditions. Administrative Report, US Geological Survey. Reston, Virginia.
Regehr, E.V., Lunn, N.J., Amstrup, S.C. and Stirling, I. 2007b. Effects of earlier sea ice breakup on survival and population size of polar bears in Western Hudson Bay. Journal ofWildlife Management 71: 2673–2683.
Reimers, E. 1982. Winter mortality and population trends of reindeer on Svalbard, Norway. Arctic and Alpine Research 14: 295–300.
Reznick, D., Bryant, M.J. and Bashey, F. 2002. r-and K-selection revisited: the role of population regulation in life-history evolution. Ecology, 83(6): 1509–1520.
Rode, K.D., Peacock, E., Taylor, M., Stirling, I., Born, E.W., Laidre, K.L. and Wiig, Ø. 2012. A tale of two polar bear populations: ice habitat, harvest and body condition. Population Ecology 54: 3–18.
Rode, K.D., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W. and Budge, S. 2013. Comparison in polar bear response to sea ice loss in the Chukchi and southern Beaufort Seas. Oral presentation at the 28th Lowell Wakefield Fisheries Symposium, March 26–29. Anchorage, AK.
Rode, K. D., Pagano, A.M., Bromaghin, J.F., Atwood, T.C., Durner, G.M. and Simac K.S. 2014a. Effects of capturing and collaring on polar bears: Findings from long-termresearch on the southern Beaufort population. Wildlife Research 41(4): 311–322.
Rode, K.D., Regehr, E.V.,Douglas,D.,Durner, G.,Derocher, A.E., Thiemann, G.W. and Budge, S. 2014b. Variation in the response of anArctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Global Change Biology 20(1): 76–88.
Schliebe, S.,Wiig, Ø., Derocher, A. and Lunn, N. (IUCN SSC Polar Bear Specialist Group) 2008. Ursus maritimus. The IUCN Red List of Threatened Species. Version 2014.3.
Smith, T.G. 1987. The Ringed Seal, Phoca hispida, of the Canadian Western Arctic. Canadian Bulletin of Fisheries and Aquatic Sciences 216. Department of Fisheries and Oceans, Ottawa.
Stirling, I. 1974. Midsummer observations on the behavior of wild polar bears (Ursus maritimus ). Canadian Journal of Zoology 52: 1191–1198.
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55 (Suppl. 1): 59–76.
Stirling, I. and Derocher, A.E. 2012. Effects of climate warming on polar bears: a review of the evidence. Global Change Biology 18(9): 2694–2706.
Stirling, I. and Lunn, N.J. 1997. Environmental fluctuations in arctic marine ecosystems as reflected by variability in reproduction of polar bears and ringed seals. In: Ecology of Arctic Environments,Woodin, S.J. and Marquiss, M. (eds). Blackwell Science.
Stirling, I. and McEwan, E.H. 1975. The caloric value of whole ringed seals (Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and hunting behaviour. Canadian Journal of Zoology 53: 1021–1027.
Stirling, I. and Parkinson, C.L. 2006. Possible effects of climate warming on selected populations of polar bears (Ursus maritimus ) in the Canadian Arctic. Arctic 59: 261–275.
Stirling, I. and Øritsland, N. A. 1995. Relationships between estimates of ringed seal (Phoca hispida ) and polar bear (Ursus maritimus ) populations in the Canadian Arctic. Canadian Journal of Fisheries andAquatic Sciences 52: 2594 – 2612.
Stirling, I., Pearson, A.M. and Bunnell, F.L. 1976. Population ecology studies of polar and grizzly bears in northern Canada. Transactions of the 41st North AmericanWildlife Conference 41: 421–430.
Stirling, I., Schweinsburg, R.E., Kolenasky, G.B., Juniper, I., Robertson, R.J. and Luttich, S. 1980. Proceedings of the 7th meeting of the Polar Bear Specialists Group IUCN/SSC, 30 January-1 February, 1979, Copenhagen, Denmark. Gland, Switzerland and Cambridge UK, IUCN, pp. 45–53.
Stirling, I, Kingsley, M. and Calvert, W. 1982. The distribution and abundance of seals in the eastern Beaufort Sea, 1974–79. Canadian Wildlife Service Occasional Paper 47. Edmonton.
Stirling, I., Lunn, N.J., Iacozza, J., Elliott, C. and Obbard, M. 2004. Polar bear distribution and abundance on the southwestern Hudson Bay coast during open water season, in relation to population trends and annual ice patterns. Arctic 57: 15–26.
Stirling, I., McDonald, T.L., Richardson, E.S. and Regehr, E.V. 2007. Polar bear population status in the Northern Beaufort Sea. Administrative Report, US Geological Survey. Reston, Virginia.
Stirling, I., Richardson, E., Thiemann, G.W. and Derocher, A.E. 2008. Unusual predation attempts of polar bears on ringed seals in the southern Beaufort Sea: possible significance of changing spring ice conditions. Arctic 61: 14–22.
Swart, N.C., Fyfe, J.C., Hawkins, E., Kay, J.E. and Jahn, A. 2015. Influence of internal variability on Arctic sea-ice trends. Nature Climate Change 5(2): 86–89.
US Fish and Wildlife Service. 2008. Determination of threatened status for the polar bear (Ursus maritimus ) throughout its range. Federal Register 73(95): 28212–28303.
US Fish and Wildlife Service. 2011. 12-month finding on a petition to list the Pacific walrus as endangered or threatened. Federal Register 76(28): 7634–7676.
US Fish and Wildlife Service. 2012a. Threatened status for the Arctic, Okhotsk and Baltic subspecies of the ringed seal. Federal Register 77(249): 76706–76738.
US Fish and Wildlife Service. 2012b. Threatened status for the Beringia and Okhotsk distinct population segments of the Erignathus barbatus nauticus subspecies of the bearded seal. Federal Register 77(249): 76740–76768.
Vibe, C. 1965. The polar bear in Greenland. In: Proceedings of the First International Scientific Meeting on the Polar Bear. Fairbanks, Alaska 6–10 September 1965, IUCN Polar Bear Specialist Group. University of Alaska International Conference Proceedings Series, No. 1. pp. 17–25. Washington, DC.
Vongraven,D., Aars, J., Amstrup, S., et al. 2012. A circumpolar monitoring framework for polar bears. Ursus 23 (sp2): 1–66.
Wang M., Overland, J.E., Stabeno, P. 2012. Future climate of the Bering and Chukchi Seas projectedby global climate models. Deep-Sea Research Part II: Topical Studies in Oceanography 65–70: 46–57.
Wu, J. and Loucks, O.L. 1995. From balance of nature to hierarchical patch dynamics: a paradigm shift in ecology. Quarterly Review of Biology 70: 439–466.
Wyatt, M.G. and Curry, J.A. 2014. Role for Eurasian Arctic shelf sea ice in a secularly varying hemispheric climate signal during the 20th century. Climate Dynamics 42: 2763–2782.
PDF of essay with foreword here.





You must be logged in to post a comment.