
Sea ice is said to be “an essential habitat for polar bears” but that’s an overly simplistic advocacy meme as ridiculous as the “no sea ice, no polar bears” message with which the public is constantly bombarded. Polar bears require sea ice from late fall to late spring only: from early summer to mid-fall, sea ice is optional. Historical evidence of polar bears that spent 5 months on land during the summer of 1874 proves an extended stay ashore is a natural response of polar bears to natural summer ice retreat, not a consequence of recent human-caused global warming. Sea ice is a seasonal requirement for polar bears: it’s not necessary year round.

[This PBI newsletter from 2011 repeats this meme and Andrew Derocher’s recent tweet conveys a similar message (“Sea ice loss = habitat loss for polar bears”)]
As long as sea ice is available from late fall through late spring (December to early June) and accompanied by abundant seal prey (sometimes it isn’t, see Derocher and Stirling 1995; Stirling 2002; Stirling et al. 1981, 1982, 1984), polar bears can survive a complete or nearly complete fast from June to late November (and pregnant females from June to early April the following year). That’s the beauty of their Arctic adaptation: fat deposited in early spring allows polar bears to survive an extraordinary fast whether they spend the time on land or sea ice.
Young and very old bears, as well as sick and injured ones, are the exception: these bears often come ashore in poor condition and end up dying of starvation — as a much-publicized bear on Baffin Island who likely had a form of cancer did last summer (Crockford 2018). Competition with bigger, stronger bears means these bears can’t keep what they are able to kill and they are most often the bears who cause problems. Starvation is the leading natural cause of death for polar bears because if they cannot put on the fat they need in spring, they will not survive the low food months of summer and winter, whether they are on land or out on the sea ice (Amstrup 2003).
Polar bear survival depends on the consumption of large numbers of fat, newborn seals that are only available in abundance from March to mid May (depending on the location and species of seal): after this time, fewer seals are available and are very hard to catch.
As I’ve stated previously (Crockford 2018:15):
“Sea ice extent in June has declined, on average, from just over 12 mkm2 in the 1980s to just over 11 mkm2 from 2004-2017. 140
By late May to early June, the young seals that form the bulk of polar bear diets in spring take to the water to feed and are no longer available on the ice, leaving only predatory-savvy adults and subadults hauled out as potential prey. 141
This means few seals are actually caught and consumed by polar bears after about mid-June in Seasonal and Divergent sea ice ecoregions, or by mid-July in Convergent and Archipelago regions (see Section 6, Prey Base).”
The most pessimistic predictions of March sea-ice extent at the end of the 21st century is about 12.0 million km2 (Stroeve et al. 2007), equal to the average extent of ice for May 2016 (shown below, from NSIDC), which is a perfectly adequate amount of ice to meet polar bear needs in all subpopulations during late winter/early spring:
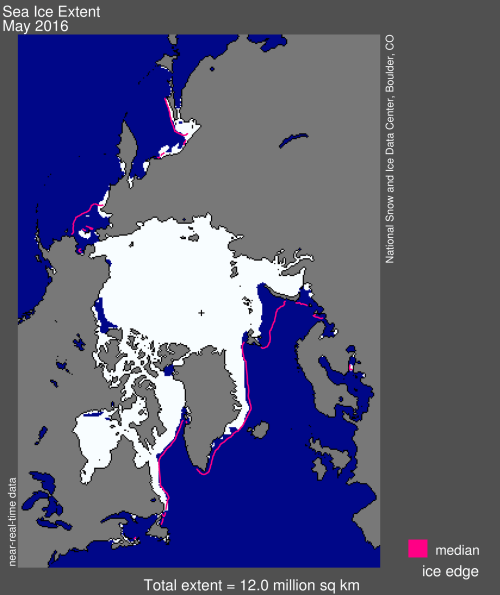
Not a single sea ice prediction suggests the disappearance of sea ice in winter or early spring due to human-caused global warming (Amstrup et al. 2007; Regehr et al. 2016) and predictions of catastrophic polar bear losses due to sea ice declines have failed to materialize (Crockford 2017).
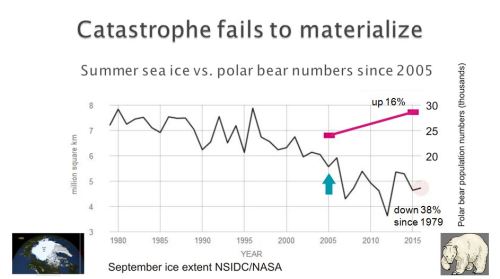
From early summer to mid-fall, many bears lounge around on land with no ill-effects. As far as we know, they have always done so (see Historical Evidence below). There is also no biological reason to suggest that well-fed bears that historically did not have to spend much time ashore in summer (such as those in the Southern Beaufort) are incapable of doing so for 5 months if necessary.
Historical Evidence
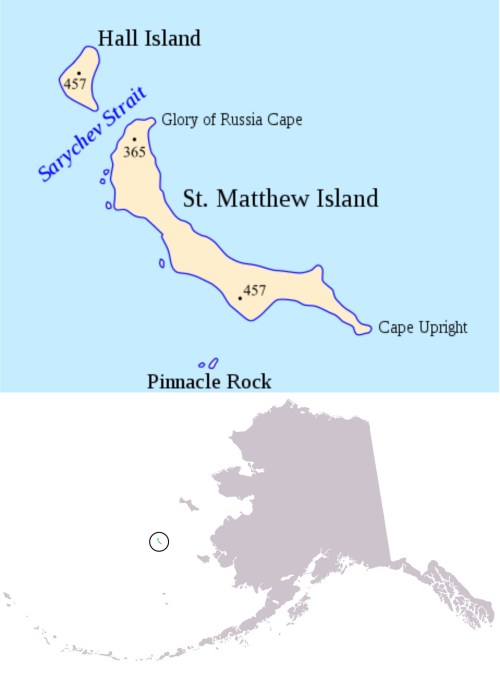
Back in 1874 — well before human-caused global warming reared its ugly head — hundreds of fat, healthy Chukchi Sea polar bears (see drawing below) spent four to five months on St. Matthew Island in the Bering Sea during the summer (at about 60°N latitude, not quite as far south as Churchill, Manitoba); some females stayed on to have their cubs in maternity dens dug into the hills (Eliott and Coues 1875; Elliott 1875; Klein and Sowls 2011).

Figure 2. A drawing of polar bears on St. Matthew Island that accompanied the May 1, 1875 Harper’s Weekly Journal of Civilization article written by Henry Elliot. See here.
From my 2013 post on this topic:
“Elliott’s government report (Elliott and Coues 1875: Appendix) notes that the bears they saw were “in most excellent condition, fat and sleek.” They encountered both males and females with twin cubs. Maynard’s report apparently mentions twins and triplets – “about one third grown” (cited in Klein and Sowls 2011:430), i.e. one year old cubs. Elliott and Maynard found empty maternity dens that had been dug into the hillsides and “bear roads” winding around the island. Fresh water was abundant on the island and while there were walrus and a few seals offshore, there was only scant evidence (the carcass of one young walrus) that the bears were feeding on them. The bears appear to have been fasting – except for the odd bit of scavenging and grass-chewing, which all land-bound polar bears appear to do.”
The fact that well-fed Chukchi Sea polar bears spent 5 months onshore in the late 1800s suggests that well-fed polar bears throughout the Arctic have always had the ability to fast for this length of time: it is not a new phenomenon associated with recent sea ice changes blamed on human-caused global warming (Overland and Wang 2013; Wang and Overland 2012, 2015).
Note that polar bears no longer den or spend the summer on St. Matthew Island because they were exterminated by commercial hunters. Wrangel Island to the north is now the primary denning/summering location.

Modern Evidence
Wrangel Island
One of the largest terrestrial denning area in the Arctic is on Wrangel Island, off the Russian coast of the Chukchi Sea, where in 2017 almost 600 bears were estimated onshore for the 3-4 month summer ice-free season. Bears in the Chukchi Sea are doing very well, better than they were in the 1980s (Rode and Regehr 2010; Rode et al. 2013, 2014, 2018), despite a dramatic reduction in summer sea ice (Serrez et al. 2016).
Last year, polar bear biologist Eric Regehr (US Fish & Wildlife Service) told the Daily Mail (23 November 2017) that the Chukchi Sea subpopulation “appears to be productive and healthy.”

Photo below of a fat Wrangel Island bear. Shutterstock.
Svalbard
Despite concerns over the effect of spring sea ice loss around the Svalbard archipelago in recent years (circled in yellow in the maps below, from Walsh et al. 2017), polar bear data collected by researchers up to the spring of 2018 show little to no negative impact from these low ice springs (see previous post here, with references).

It must be remembered that the polar bear subpopulation region that encompasses Svalbard is called “Barents Sea” (see map below from the PBSG) and includes the archipelago of Franz Josef Land to the east that has a much colder climate (Barr 1995). Franz Josef Land is where most Barents Sea polar bears live (Aars et al. 2009) and provides abundant denning habitat for pregnant females as well as a refugium for bears that prefer to stay on land during the summer when sea ice retreats. It is likely that most females that formerly made terrestrial dens on Svalbard have now shifted to Franz Josef Land (Aars 2015; Aars et al. 2017; Descamps et al. 2017), except for years (like 2014) with abundant fall ice.

Western and Southern Hudson Bay
Here are critical words to remember (more details here) from biologist Martin Obbard and colleagues (2016:29) on the relationship between body condition and sea ice for Southern Hudson Bay (SH) polar bears, which apply equally well to bears in other regions:
“Date of freeze-up had a stronger influence on subsequent body condition than date of break-up in our study. Though models with date of freeze-up were supported over models with other ice covariates, we acknowledge that lower variability in freeze-up dates than in ice duration or break-up dates could have influenced the model selection process. Nevertheless, we suggest that a stronger effect of date of freeze-up may be because even though break-up has advanced by up to 3-4 weeks in portions of Hudson Bay it still occurs no earlier than late June or early July so does not yet interfere with opportunities to feed on neonate ringed seal pups that are born in March-April in eastern Hudson Bay (Chambellant 2010). Therefore, losing days or weeks of hunting opportunities during June and July deprives polar bears of the opportunity to feed on adult seals, but does not deprive them of the critical spring period (Watts and Hansen 1987) when they are truly hyperphagic. No doubt, the loss of hunting opportunities to kill adult seals has a negative effect on body condition, but it appears that for bears in SH a forced extension of the fast in late fall has a greater negative effect on subsequent body condition.” [my bold]
In other words, by mid-June at least, polar bears have largely finished their intensive feeding that’s so critical to their survival over the rest of the year. They may catch a few seals over the coming months but for most bears, this makes little difference to their overall condition or potential survival.
Most bears are at their fattest in early summer (when they come off the ice to spend the summer ashore) after having gorged on newborn seals in early spring.
Breakup dates for Western Hudson Bay have not become progressively earlier each year since 1979: rather, a step-change occurred about 1997/1998 that meant breakup dates since then have been about 3 weeks earlier than before (with much year to year variation). There has been no trend in breakup or freeze-up dates since 1995 or 2001 depending on how you calculate the data (Castro de la Guardia et al. 2017; Lunn et al. 2016).
For both Western and Southern Hudson Bay, very late freeze up has had the most negative impact on polar bear survival. When bears come ashore in less than good condition (as they did in 1983), many bears can struggle to survive. However, poor feeding conditions on the bay during early spring (about which virtually nothing is known), can also impact the body condition and survival of bears.
References
Aars, J. 2015. Research on polar bears at Norwegian Polar Institute. Online seminar (‘webinar”), January 14. pdf here.
Aars, J., Marques,T.A, Lone, K., Anderson, M., Wiig, Ø., Fløystad, I.M.B., Hagen, S.B. and Buckland, S.T. 2017. The number and distribution of polar bears in the western Barents Sea. Polar Research 36:1. 1374125. doi:10.1080/17518369.2017.1374125
Aars, J., Marques, T.A., Buckland, S.T., Andersen, M., Belikov, S., Boltunov, A., et al. 2009. Estimating the Barents Sea polar bear subpopulation. Marine Mammal Science 25: 35-52.
Amstrup, S.C. 2003. Polar bear (Ursus maritimus). In Wild Mammals of North America, G.A. Feldhamer, B.C. Thompson and J.A. Chapman (eds), pg. 587-610. Johns Hopkins University Press, Baltimore.
Amstrup, S.C., Marcot, B.G. & Douglas, D.C. 2007. Forecasting the rangewide status of polar bears at selected times in the 21st century. US Geological Survey. Reston, VA. Pdf here
Barr, S. 1995. Franz Josef Land. Oslo: Norwegian Polar Institute. ISBN
82-7666-095-9.
Castro de la Guardia, L., Myers, P.G., Derocher, A.E., Lunn, N.J., Terwisscha van Scheltinga, A.D. 2017. Sea ice cycle in western Hudson Bay, Canada, from a polar bear perspective. Marine Ecology Progress Series 564: 225–233. http://www.int-res.com/abstracts/meps/v564/p225-233/
Crockford, S.J. 2017. Testing the hypothesis that routine sea ice coverage of 3-5 mkm2 results in a greater than 30% decline in population size of polar bears (Ursus maritimus). PeerJ Preprints 2 March 2017. Doi: 10.7287/peerj.preprints.2737v3 Open access. https://doi.org/10.7287/peerj.preprints.2737v3
Crockford, S.J. 2018. State of the Polar Bear Report 2017. Global Warming Policy Foundation Report #29. London. pdf here.
Derocher, A.E. and Stirling, I. 1995. Temporal variation in reproduction and body mass of polar bears in western Hudson Bay. Canadian Journal of Zoology 73:1657-1665. http://www.nrcresearchpress.com/doi/abs/10.1139/z95-197
Descamps, S., Aars, J., Fuglei, E., Kovacs, K.M., Lydersen, C., Pavlova, O., Pedersen, Å.Ø., Ravolainen, V. and Strøm, H. 2017. Climate change impacts on wildlife in a High Arctic archipelago — Svalbard, Norway. Global Change Biology 23: 490-502. doi: 10.1111/gcb.13381
Elliott, H.W. 1875 . Polar bears on St. Matthew Island. Harper’s Weekly Journal of Civilization. May 1 issue. Harper and Brothers, New York.
Elliott, H.W. and Coues, E. 1875. A report upon the condition of affairs in the territory of Alaska. US Government Printing Office, Washington. http://tinyurl.com/a8zk6yk
Klein, D.R. and Sowls, A. 2011. History of polar bears as summer residents on the St. Matthew Islands, Bering Sea. Arctic 64:429-436. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/4142
Lunn, N.J., Servanty, S., Regehr, E.V., Converse, S.J., Richardson, E. and Stirling, I. 2016. Demography of an apex predator at the edge of its range – impacts of changing sea ice on polar bears in Hudson Bay. Ecological Applications, in press. DOI: 10.1890/15-1256
Obbard, M.E., Cattet, M.R.I., Howe, E.J., Middel, K.R., Newton, E.J., Kolenosky, G.B., Abraham, K.F. and Greenwood, C.J. 2016. Trends in body condition in polar bears (Ursus maritimus) from the Southern Hudson Bay subpopulation in relation to changes in sea ice. Arctic Science, in press. 10.1139/AS-2015-0027
Overland, J.E. and Wang, M. 2013. When will the summer Arctic be nearly sea ice free? Geophysical Research Letters 40: 2097-2101.
Regehr, E.V., Laidre, K.L, Akçakaya, H.R., Amstrup, S.C., Atwood, T.C., Lunn, N.J., Obbard, M., Stern, H., Thiemann, G.W., & Wiig, Ø. 2016. Conservation status of polar bears (Ursus maritimus) in relation to projected sea-ice declines. Biology Letters 12: 20160556. http://rsbl.royalsocietypublishing.org/content/12/12/20160556 Supplementary data here.
Rode, K. and Regehr, E.V. 2010. Polar bear research in the Chukchi and Bering Seas: A synopsis of 2010 field work. Unpublished report to the US Fish and Wildlife Service, Department of the Interior, Anchorage. pdf here.
Rode, K.D., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2013. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Oral presentation by Karyn Rode, 28th Lowell Wakefield Fisheries Symposium, March 26-29. Anchorage, AK.
Rode, K.D., Regehr, E.V., Douglas, D., Durner, G., Derocher, A.E., Thiemann, G.W., and Budge, S. 2014. Variation in the response of an Arctic top predator experiencing habitat loss: feeding and reproductive ecology of two polar bear populations. Global Change Biology 20(1):76-88. http://onlinelibrary.wiley.com/doi/10.1111/gcb.12339/abstract
Rode, K. D., R. R. Wilson, D. C. Douglas, V. Muhlenbruch, T.C. Atwood, E. V. Regehr, E.S. Richardson, N.W. Pilfold, A.E. Derocher, G.M Durner, I. Stirling, S.C. Amstrup, M. S. Martin, A.M. Pagano, and K. Simac. 2018. Spring fasting behavior in a marine apex predator provides an index of ecosystem productivity. Global Change Biology http://onlinelibrary.wiley.com/doi/10.1111/gcb.13933/full
Serreze, M.C., Crawford, A., Stroeve, J.C., Barrett, A.P. and Woodgate, R.A. 2016. Variability, trends and predictability of seasonal sea ice retreat and advance in the Chukchi Sea. Journal of Geophysical Research 121 (10):7308–7325. http://onlinelibrary.wiley.com/doi/10.1002/2016JC011977/abstract
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55 (Suppl. 1):59-76. http://arctic.synergiesprairies.ca/arctic/index.php/arctic/issue/view/42
Stirling, I., Andriashek, D., and Calvert, W. 1993. Habitat preferences of polar bears in the western Canadian Arctic in late winter and spring. Polar Record 29:13-24. http://tinyurl.com/qxt33wj
Stirling, I., Calvert, W., and Andriashek, D. 1984. Polar bear ecology and environmental considerations in the Canadian High Arctic. Pg. 201-222. In Olson, R., Geddes, F. and Hastings, R. (eds.). Northern Ecology and Resource Management. University of Alberta Press, Edmonton.
Stirling, I, Cleator, H. and Smith, T.G. 1981. Marine mammals. In: Polynyas in the Canadian Arctic, Stirling, I. and Cleator, H. (eds), pg. 45-58. Canadian Wildlife Service Occasional Paper No. 45. Ottawa.
Stirling, I, Kingsley, M. and Calvert, W. 1982. The distribution and abundance of seals in the eastern Beaufort Sea, 1974–79. Canadian Wildlife Service Occasional Paper 47. Edmonton.
Stroeve, J., Holland, M.M., Meier, W., Scambos, T. and Serreze, M. 2007. Arctic sea ice decline: Faster than forecast. Geophysical Research Letters 34:L09501.
Walsh, J.E., Fetterer, F., Stewart, J.S. and Chapman, W.L. 2017. A database for depicting Arctic sea ice variations back to 1850. Geographical Review 107(1):89-107. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1931-0846.2016.12195.x
Wang, M. and Overland, J.E. 2012. A sea ice free summer Arctic within 30 years: An update from CMIP5 models. Geophysical Research Letters 39:L18501.
Wang, M. and Overland, J.E. 2015. Projected future duration of the sea-ice-free season in the Alaskan Arctic. Progress in Oceanography 136:50-59.





You must be logged in to post a comment.